All About Archaeopteryx
by Chris Nedin[References Edited: June 10, 2002]
![]()
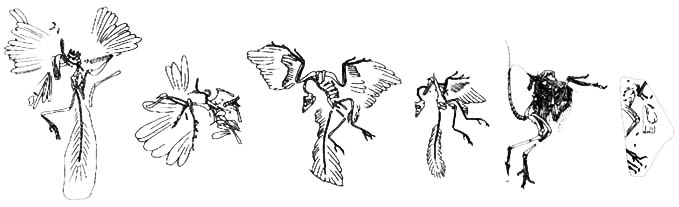
Contents
- Introduction
- Archaeopteryx specimens
- Archaeopteryx features
- Archaeopteryx + modern birds, just variation within kind?
- Ratites
- A tale of two pelvises
- To fly or not to fly
- Archaeopteryx's Ancestry
- Protoavis
- Conclusions
- Acknowledgements
- References
Introduction
Archaeopteryx lithographica ("ancient wing from the printing stone").
Named after the limestone in which it was discovered. The stone is a smooth, fine grained limestone which was used in printing. Quarried from in and around the Solnhofen area of Germany. Formed on the bottom of a hypersaline lagoon in the Late Jurassic, about 150 million years ago.
Archaeopteryx specimens
There have been 8 specimens of Archaeopteryx found (7 actual specimens and one feather). These finds are documented chronologically (by description) below.
- The Feather
Found in 1860 near Solnhofen and a revelation when it was described by H. v Meyer in 1861. The surprise was not the age of the fossil, since several ornithopod dinosaur footprints erroneously ascribed to birds were known from the Triassic, but the detail that was preserved.
- The London Specimen

Found in 1861, near Langenaltheim. Probably the best known (together with the Berlin specimen). Its discovery was announced by H. v Meyer in 1861 and the specimen was subsequently bought by the British Museum of Natural History in London (under the instruction of Richard Owen). It cost 700 UK Pounds - a small fortune in those times, but for that price Owen also received just over a thousand other fossils from Solnhofen. The specimen was sold by amateur collector and local doctor Carl Haberlein, who had received it in lieu of payment for medical treatment. Owen described the specimen in 1863. He saw at once that it was an important find and recognised that it represented a transitional form - but not in the "Darwin" sense. Owen was a staunch "evolutionist", however he did not believe in Darwin's model of evolution. Interestingly Huxley, who was a staunch "Darwinist" failed to recognise the true import of the fossil and merely remarked on it as a "reptile-like bird". It wasn't until close comparisons were made with the dinosaur Compsognathus that Archae's true worth was realised. - The Berlin Specimen

Found in 1877 near Blumenberg. This was a better specimen than the London specimen, principally because it had a complete head, albeit badly crushed, and was snapped up by the Berlin museum. It was sold to them by Carl Haberlein's son (talk about keeping it in the family!). It was described by W. Dames in 1884.
- The Maxburg Specimen

Found in 1958 near Langenaltheim (same as London Specimen). This specimen is of the torso only and is the only specimen to still be in private hands. In 1992, after the death of its finder and owner Eduard Opitsch, the specimen was found to be missing and it is thought that it was sold secretly (Abbott 1992). It's whereabouts remain unknown. The specimen was described by Heller in 1959.
- The Haarlem or Teyler Specimen

This specimen was actually found near Reidenburg in 1855, 5 years before the feather! It lay in a museum after being classified as Pterodactylus crassipes by H. v Meyer in 1875. Curiously, Mayer described it as having a flight membrane unlike any other known pterodactyl, now we know why! A re-examination of the fossil in 1970 by Ostrom revealed feathers and its true identity.
- The Eichstatt Specimen

Found near Workerszell in 1951, it was described by P. Wellnhofer in 1974. This is the smallest of all the specimens, being some 2/3 the size of the others. It also differs in other aspects such as the tooth structure and the poorly ossified shoulder bones. It has been suggested that this is a separate genus, however the differences can also be ascribed to the possible juvenile stage of the animal and/or a different feeding niche. However, this specimen has the best preserved head, from which the litany of Archae's reptilian cranial features were described. At the moment it still resides within A. lithographica.
- The Solnhofen Specimen

Found in the 1960's near Eichstatt by a Turkish worker. First identified as Compsognathus, by a amateur collector, however, further examination showed that the arms were too long for the body size and preparation revealed feather traces. Described by P. Wellnhofer in 1988.
- The Solnhofen-Aktien-Verein specimen
A new specimen was described by Wellnhofer (1993), but the description is in German and so information is limited. The specimen has been classified as a new species, Archaeopteryx bavarica, and has been reported as possessing a small ossified sternum, as well as feather impressions.
Archaeopteryx Features
Much has been made in pseudoscientific circles about the position of Archae within the evolutionary scheme of things. The usual "argument" put forward is that Archae cannot be a transitional fossil between birds and dinosaurs because it is a bird. This simplistic line belies the fact that, whilst Archae is indeed classified as a bird, it has been done so on the strength of 4 main characters - 2 of which are not unique to birds. This classification ignores the fact that Archae has numerous characters which are unique, unique in that they are not possessed by birds. Archae's avian affinities are allowable on the strength of the following 4 main characters:
Archaeopteryx's avian features
- 1) Feathers.
-
Feathers are the diagnostic feature of modern birds. This is one of the main criterion for classifying Archae as a bird, as no other modern animal has feathers. The possession of feathers is a characteristic of birds, so strike one up for the birds. However, in late 1996, a discovery in China may change this view. A small theropod dinosaur Sinosauropteryx (Chen et al. 1998) was found with what appear to be feathers preserved along the back. The identification of the sturctures is equivocal however, (e.g. Unwin 1998), with some doubting that the structures are feathers.
STOP PRESS
Feathered Dinosaurs Found
Two species of dinosaur have recently been found in northeast China which possess feathers (Qiang et al. 1998). Protoarchaeopteryx robusta and Caudipteryx zoui show regiges, rectrices and plumulaceous feather inpressions. Further, they are not birds, lacking a reverted (backwards facing) big toe (see number 2 below) and a quadrratojugal squamosal contact, having a quadrojugal joined to the quatrate by a ligament and a reduced or absent process of the ishium. These and other characters group Protoarchaeopteryx and Caudipteryx with maniraptoran coelurosaurs rather than birds.
[Systematics Note (from Padian 1998): Systematists define the names of organisms by their ancestry, in this case birds (Aves) consist of Archaeopteryx plus living birds and all the descendants of their most recent common ancester. Birds are diagnosed by unique features that only they possess and which are inherited from that common ancester. Even if feathers are shared by a wider group than just birds, birds are still defined as Archaeopteryx and later relatives. Protoarchaeopteryx and Caudipteryx are not birds even though they have feathers because the suite of morphological characters they possess mark them as belong to the maniraptoran coelurosaur dinosarus.]
It appears that feathers can no longer be used as a unique feature of birds.
For a discussion of the feathered dinosaurs go to Jeff Poling's Dinosauria page.
- 2) Opposable hallux (big toe).
-
This also is a character of birds and not of dinosaurs. Although opposable big toes are found in other groups, they are not, as far as I am aware, found in dinosaurs. A reversed big toe is found in some dinosaurs however, and the condition is approached in some theropod dinosaurs.
- 3) Furcula (wishbone) formed of two clavicles fused together in the midline.
-
Now we start getting on shaky ground. It used to be thought that the possession of a furcula distinguished birds from dinosaurs. Indeed, up until recently even clavicles were few and far between in even theropod dinosaurs (the suggested closest group to the birds and from which the birds evolved - see Ostrom 1976). However, it has been found that theropod dinosaurs did indeed have clavicles (e.g. Bryant & Russell 1993) and they have been found in several species, e.g., Segisaurus, Velociraptor, Euparkeria, Ornithosuchus, Saltoposuchus, Ticinosuchus. Also, Chure & Madson (1996) reported furculae in a non-maniraptoran, allosaurid dinosaur.
It has been found that the clavicles are often small and poorly ossified. This is no surprise, since they are of little evolutionary advantage to your average theropod dinosaur. However, birds too show this variation in ossification, especially amongst the carniates and some parrots, clavicles are reduced or even missing. Therefore the apparent absence of clavicles in some theropod dinosaurs may well be due to poor ossification rather than true absence. However, furculas have been found in some theropod dinosaurs, namely the Oviraptorosauria (Barsbold et al. 1990, Bryant & Russell 1993), for example Oviraptor and Ingenia. Thus furculas do not appear to be diagnostic to birds and certain members of the suggested closest group to the birds now appear to possess furculas so it is a neutral character.
A commonly cited criticism of this is that most of the theropod dinosaurs listed here post-date Archae. However, none of these is claimed as the ancestor anyway, and Eupakeria is a Triassic form. The presence of clavicles shows that this character is a feature of theropod dinosaurs and thus was probably present in early theropods.
- 4) Pubis elongate and directed backward.
-
This is a feature of birds, but it is also a feature of some theropod dinosaurs so is not diagnostic of birds - another neutral character. However, the pubic shafts of Archaeopteryx and dromaeosaurs (a group of theropod dinosaurs which are thought to be closely linked to birds) share a plate-like, slightly angled transverse cross-section which not found in any other archosaurs.
Archaeopteryx's reptile features
- 5) Premaxilla and maxilla are not horn-covered.
-
This is posh talk for "does not have a bill." The premaxilla does not have a keratinized covering, so Archaeopteryx has no bill. The bill is produced via the process of 'cornification' which involves the mucus layer of the epidermis (Romanoff 1960) and thus its formation is independant of jaw bone formation.
- 6) Trunk region vertebra are free.
-
In birds the trunk vertebrae are always fused.
- 7) Bones are pneumatic.
-
I.e. they appear to have air-sacs, as they do in birds and in some dinosaurs (e.g. Witmer 1990, Brooks 1993). It should be pointed out that previous claims suggesting the bones of Archae were not pneumatic (Lambrecht 1933; de Beer 1954), was based on negative evidence, i.e. that the bones do not exhibit pneumatic pores (through which the air sacs enter the bones) and the bones show none of the plumpness and bulges which characterise the pneumatic bones of modern birds. Britt et al. (1998) found evidence for the presence of pnematic bones in Archaeopteryx:
"Here we re-examine two specimens of _Archaeopteryx_. These specimens show evidence of vertebral pneumaticity in the cervical and anterior thorasic vertebrae, thus confirming the phylogenetic continuity between the pneumatic systems of non-avialan theropods and living birds" (Britt et al. 1998, p. 374)
8) Pubic shafts with a plate-like, and slightly angled transverse cross-section-
A Character shared with dromaeosaurs but not with other dinosaurs or birds
- 9) Cerebral hemispheres elongate, slender and cerebellum is situated behind the mid-brain and doesn't overlap it from behind or press down on it.
-
This again is a reptilian feature. In birds the cerebral hemispheres are stout, cerebellum is so much enlarged that it spreads forwards over the mid-brain and compresses it downwards. Thus the shape of the brain is not like that of modern birds, but rather an intermediate stage between dinosaurs and birds (e.g. Alexander 1990).
- 10) Neck attaches to skull from the rear as in dinosaurs not from below as in modern birds.
- The site of neck attachement (from below) is
characteristic in birds, _Archaeopteryx_ does not have this
character, but is the same as theropod dinosaurs:
"Notice that this coelurosaurian-like neck extended back from the rear of the skull in _Archaeopteryx_ - as it does in coelurosaurs [theropod dinosaurs], rather than from beneath as in later birds." (Ostrom 1976, p. 137).
Skull and brain of Archae is basically reptilian and is not "totally birdlike" (contrary to a certain creationist's claim).
- 11) Center of cervical vertebrae have simple concave articular facets.
-
This is the same as the archosaur pattern. In birds the vertebrae are different, they have a saddle-shaped surface:
"The most striking feature of the vertebrae is the simple disk-like facets of their centra, without any sign of the saddle-shaped articulations found in other birds" (de Beer 1954, p. 17).
- 12) Long bony tail with many free vertebrae up to tip (no pygostyle).
-
Birds have a short tail and the caudal vertebrae are fused to give the pygostyle.
- 13) Premaxilla and maxilla bones bear teeth.
-
No modern bird possess teeth (e.g. Romanoff 1960; Orr 1966, p. 113). Bird embryos form tooth buds, but do not actually produce teeth. Some birds subsequently produce ridges in the bill, but there is no connection between them and the embryonic tooth buds, since the ridges also form in other areas of the bill where no tooth buds have previouslu formed. Some birds produce hook-like structures which are papillae, and appear to be related to the process of keratinization of the beak (Romanoff 1960), and have nothing to do with teeth. They do not possess blood vessel or nerve connections, nor do they produce dentine.
The expression of tooth buds in the bird embryo has a simple evolutionary explanation, since it suggests that the ancestors of modern birds possessed teeth and that this character has been supressed in modern birds. The presence of tooth buds in the embryos of organisms which do not possess teeth in the adult is a difficulty for anti-evolutionists, since why should a character be expressed that is never used in the organism? Some fossil birds exhibit a reduction in the number of bones which have teeth. Both Hesperornis and Baptornis lack teeth on the premaxilla (Archaeopteryx and theropod dinosaurs have teeth on both the maxilla and premaxilla). Not only that, Hesperornis has a beak, but on the upper jaw only (Gingerich 1975). It therefore has half a beak and teeth. A good example of a morphologicaly intermediate structure between toothed birds which lack a beak, and beaked, toothless birds.
- 14) Ribs slender, without joints or uncinate processes and do not articulate with the sternum.
-
Birds have stout ribs with uncinate processes (braces between them) and articulate with the sternum.
- 15) Pelvic girdle and femur joint is archosaurian rather than avian (except for the backward pointing pubis as mentioned above).
-
Here Archae really shows its transitional nature. Whilst the pelvic girdle as a whole is basically free and similar to archosaur girdles, the pubis points backward - a character shared with birds and some other bird-like theropod dinosaurs.
What is interesting is that with the bird pelvis:
"The ischium lies beneath the posterior part of the ilium and beneath this again is the pubis, which is directed backwards (i.e. like this: =). Embryological studies show that the peculiar position of these bones is the result of secondary rotation and that the pectineal process, in front of the ascetabulum, is not the true pubis as some workers have maintained." (Bellairs & Jenkin 1960, p. 258).
In other words, the embyonic pelvis of the bird, when first formed, looks, in shape and angle between the ilium and the pubis (45 degrees), very similar to the "A"-frame pelvis of Archaeopteryx (i.e. like this: <) (e.g. Romanoff 1960). The fully formed pelvis with all bones lying parallel is the result of secondary rotation of the pubis from "<" to "=". This supports the view that birds had an ancester with a saurischian pelvis such as the type possessed by Archaeopteryx and other theropod dinosaurs. (see also A tale of two pelvises below)
- 16) The Sacrum (the vertebrae developed for the attachment of pelvic girdle) occupies 6 vertebra.
-
This is the same as in reptiles and especially ornithipod dinosaurs. The bird sacrum covers between 11-23 vertebrae! So, while the variation seen in modern birds is large, it is nowhere near the number found in Archaeopteryx
- 17) Metacarpals (hand) free (except 3rd metacarpal), wrist hand joint flexible.
-
This is as in reptiles. In birds the metacarpals are fused together with the distal carpals in the carpo-metacarpus, wrist /hand fused. All modern birds have a carpo-metacarpus, all fossil birds have a carpo-metacarpus - except one (guess!) :-). However, the carpals of several coelurosaur dinosaur groups show a trend towards fusion, and in the Late Cretaceous form Avimimus, a true carpo- metacarpus is formed.
It has been suggested that the ostrich and/or other Ratites also possess unfused wrist/hand bones. This is not correct:
"The ostrich, emus, rheas, cassowaries and kiwis are often referred to together as the Ratites, though they may not be closely related to each other. They have tiny wings and cannot fly, but the bones of their hands are fused together in the same peculiar way as in flying birds, which suggest that they evolved from flying birds." (Alexander 1990, p. 435).
Some similarity between the hand of the ostrich and some of the more derived theropod dinosaurs was once used to suggest that the Ratites were 'primitive' and evolved before the advent of flight in birds. However Tucker (1938b) showed that such similarities are entirely superficial.
"He has directed attention to the bird-like characters of the hand of the dinosaur Ornitholestes as evidence that a bird-like hand can be developed independantly of flight, but the writer has pointed out in the communication mentioned above [Tucker 1938b] that the resemblance is utterly superficial and that the peculiar bowing and terminal fusion of metacarpals 2 and 3 which charcaterise both the Carnate and the Ratite hand are in no wise [sic?] reproduced in the dinosaur." (Tucker 1938a, p. 334).
"Reverting now to the reasons on which have sought to base the view that the Ratites were primitive birds whose ancesters had never flown, one: the similarity between the hand of the ostrich and that of the dinosaur, has been dismissed as invalid. Tucker (1938b) has shown that such resemblances as there are between them are only superficial and without significance." (de Beer 1956, p. 65).
- 18) Nasal opening far forward, separated from the eye by a large preorbital fenestra (hole).
-
This is typical of reptiles, but not of birds. Where a fenestra is present in birds, it is always greatly reduced, and is involved in prokinesis (movement of the beak)
- 19) Deltoid ridge of the humerus faces anteriorly as do the radial and ulnar condyles.
-
Typical of reptiles but not found in birds
- 20) Claws on 3 unfused digits.
-
No modern adult bird has 3 claws, nor do they have unfused digits. The juvenile hoatzin and Touracos do have 2 claws but loose them as they grow, the ostrich appears to retain its 2 claws into adulthood, due to the early termination of development (see section on Ratites). In the case of the hoatzin it is thought that these claws allow the juvenile to climb. It had been claimed that since these birds do have claws, even in the juvenile stage, then the presence of claws cannot be used as a reptilian character. This is not so, however. In fact almost all birds exhibit claws, but in the embryonic stage and they are lost by the time the bird leaves the egg. In the case of the few which do retain claws into the juvenile stage, this is merely the extension of the condition into the post-embryonic stage. As McGowan (1984, p 123) says:
"In retaining a primitive reptilian feature which other birds lose just before leaving the egg [the hoatzin] is showing us its reptilian pedigree. Far from being evidence to the contrary, the hoatzin is additional evidence for the reptilian ancestry of birds."
- 21) The fibula is equal in length to the tibia in the leg.
-
This again is a typical character of reptiles. In birds the fibula is shortened and reduced.
- 22) Metatarsals (foot bones) free.
-
In birds these are fused to form the tarsometatarsus. However, in modern bird embryos, the foot bones are initially separate as in the adult Archaeopteryx and is another character supporting a reptilian ancestry for birds. After all, why bother producing separate bones in the embryo and then fuse them? Why not produce a fused mass to start with? No adult modern bird has separate metatarsals, but they are separated, initially, in the embryo. This can be explained in terms of evolution - birds evolved from a group which had unfused metatarsals.
Ceratosaurians, Avimimus, and Elmisauridae all show true tarso-metatarsi. Archae itself only shows the beginning of this structure.
- 23) Gastralia present.
-
Gastralia are "ventral ribs," elements of dermal bone in the ventral wall of the abdomen. Typical of reptiles, they are absent in birds, e.g.:
"In addition to the true ribs the British Museum specimen shows a large number of so-called ventral ribs or gastralia, elements of dermal bone lying in the ventral wall of the abdomen." (de Beer 1954, p. 18)
"The gastralia of the Berlin specimen are identical with those of the British Museum specimen, but more have been preserved." (de Beer 1954, p. 19)
"The "new" specimen was found 8 September 1970 on display in the Teyler Museum, Haarlem, Netherlands. It consists of two small slabs (specimens 6928 & 6929), part and counterpart which contain impressions or parts of the left manus and forearm, pelvis, both legs and feet, and some gastralia." (Ostrom 1970, p. 538)
"Also present are numerous fragments of gastralia, faint impressions of three or four dorsal vertebrae, . . " (Ostrom 1972, p. 291).
"The counterpart slab (No. 6929) contains additional gastralia, phalanges, .." (Ostrom 1972, p. 291)
"Gastralia, or dermal abdominal ribs are present in all five skeletal specimens of _Archaeopteryx_" (Ostrom 1976, p. 139-140).
Gastralia are present on the Eichstatt specimen (See Wellnhofer 1974, fig. 7C)
Table of archaeopteryx features
1 = present; * = present in some; ? = possibly
present; x = absent
Dinosaurs Archae Birds
1 * 1 1
2 x 1 1
3 * 1 1
4 * 1 1
5 x x 1
6 x x 1
7 * x 1
8 * 1 x
9 1 1 x
10 1 1 x
11 1 1 x
12 1 1 x
13 1 1 x
14 1 1 x
15 1 1 x
16 6 6 11-23
17 1 1 x
18 1 1 *
19 1 1 x
20 1 1 x
21 1 1 x
22 1 1 x
23 1 1 x
It can be seen that Archae possesses many more characters which are present in dinosaurs and not in birds, than it does characters which are present in birds but not in dinosaurs. This is why Archae is a true transitional species, because it shares some characters which are diagnostic of one group whilst still retaining characters diagnostic of its ancestral group. Anyone who claims that Archae is 100% bird is wrong. Anyone who claims that Archae's skeleton is even predominantly bird- like is wrong. Anyone who claims Archae has a "totally birdlike" skull is wrong.
This latter point is made in reference to the claim by Dr. Duane Gish that the skull of Archae is "totally birdlike" (R. Trott pers. comm. 1994). This claim is false. To show this we need to consider the skull of Archae further.
Cranial features of Archaeopteryx
As stated above, Dr Gish claims that the skull of Archae is "totally birdlike." This is false. Romer (1950 p. 261) describes Archae thus: "The skull, as far as can be seen, was rather birdlike. . . ". However, not only is this a far cry from "totally birdlike," but Romer was using the detailed reconstruction of the Berlin Specimen, by Heilmann (1926). Ostrom (1976 p. 131) has this to say on the Heilmann reconstruction:
"Despite the details shown there [Heilmann's reconstruction of the skull -cn], the actual specimen does not permit such detailed and precise conclusions. It [the Berlin specimen's skull - cn] is badly crushed and the bones are extensively fracture, chipped and distorted - to the extent that very few cranial or mandibular sutures are unmistakably identifiable. Heilmann's reconstructions have been republished by many authors and subsequent interpretations and hypotheses based on it. Quite probably, some authors have been unaware of the inadequate basis of Heilmann's reconstruction, and understandably so unless they have had the opportunity to examine the specimen itself."
And again on the same page:
"Fortunately, the Eichstatt speciman now provides a comparative basis for evaluating and correcting past reconstructions of the Berlin skull."
As mentioned above, the Eichstatt specimen was not described until 1974, therefore Romer's description was based on the Heilmann reconstruction.
Using both specimens, Ostrom (1976 p. 132) delineated 10 characters on the skull of Archae which it shares with other theropod dinosaurs such as Ornitholestes, Compsognathus, Velociraptor and perhaps Saurornithoides. These are:
- A sharply tapered snout. In birds the maxilla makes up
the "snout". Although the snout is shortened in
Archae.
- Long elliptical external nares [nostrils] bounded
almost exclusively above and below by the premaxilla and
nasal (a bone at the front of the upper jaw). In birds the
nares are reduced and removed posteriorly (closer to the
eyes) due to the increase in size of the premaxilla.
- A large preorbital fossa and a large triangular
postorbital fossa which contains two small openings.(These
are holes in the skull. One is a single opening in front of
the orbit or eye socket, the other is behind the eye socket
and has two openings). In other fossil birds the preorbital
is reduced, the postorbital has only a single opening. In
modern birds the preorbital is greatly reduced and the
postorbital has a single opening.
- A slender, nearly vertical preorbital bar separating
the antorbital fossa and the orbit (There's that hole
again! This means that there is a vertical bar of bone
separating the hole in front of the eye with the eye
itself). Fossil and modern birds have a much more robust
preorbital bar - due to the reduction of the preorbital
fossa.
- A large circular orbit which contains a large sclerotic
ring.(The sclerotic ring is common to reptiles, birds and
actinoptygian fish, but most fossil reptiles and all fossil
birds have them. They are a series of ossified plates which
circle the eye).
- A thin, straight jugal bone makes up the zygomatic arch
(this is the bone that runs under the eye, the cheek-bone
as it extends back towards your ear, in humans). In modern
birds the arch is composed of the quadratojugal, jugal and
maxilla bones.
- A stout quadrate of moderate length which is inclined
forward. This is the bone in the upper jaw which forms part
of the jaw articulation - with the articular - in reptiles,
and Archae has a big one, um if you see what I mean!
(Incidentally - for those of you who are still with us, the
quadrate is attached to the stapes in the upper jaw, and as
we all know, the stapes is the bone which vibrates in the
ear so that we can all hear. Thus the stapes and quadrate
were attached in reptiles and it is not a great leap
forward to have both the stapes and the quadrate in the ear
as it is in mammals. Thus the claim made by Gish that - in
order for the bones to enter the ear in the transition from
reptiles to mammals they would have had to gone through a
stage whereby the 'mammal' would have deafened itself every
time it opened its mouth - is not accurate, since the
condition of stapes+quadrate articulating in the jaw joint
is found in fossil and extant reptiles, but I
digress.)
- A lower jaw which is unusually shallow and has a
conspicuous bend behind the tooth row. Not in birds.
- A long retroarticular process. Again a classic reptile
jaw-joint feature, but shared with birds.
- Premaxilla, lacrimal and jugal bones separate. In birds
the premaxilla bone is connected to the, jugal bone; the
jugal bone is connected to the, lacrimal bone. . .
:-)
- A single vomer.
- Strong curvature of the ectopterygoid hook.
- A dorsal recess in the prootic.
Although Archaeopteryx does have certain skull characters which are avian, these are insufficiant to allow the characterization "totally birdlike".
Heilmann described an external mandibular fenestra (a hole in the lower jaw) bordered below by the dentary bone - which is the condition in birds - rather than by the angular bone - as it is in reptiles. This would indicate that Archae has a lower jaw which contained both avian (said mandibular fenestra) and reptilian features (toothed lower jaw). This is all very well and dandy and is a feature which one would like to see in Archae. However, as Ostrom (1976 p. 132) points out:
"Much as I would like to accept this interpretation, the highly fractured condition of the lower jaw bone (or bones) that border the supposed mandibular fenestra, either below or above, make it impossible to certify their identifications. In fact, the fractured upper margins of the supposed fenestra leave considerable doubt as to the very existence of a 'fenestra' - a doubt which has not been removed by the Eichstatt specimen." Thus, far from conspiring to present Archae in the best possible light, a very useful feature which would have aided in the description of Archae as a true transitional fossil has in fact been show to be, in all probability, not true. So much for the evilutionist conspiracy! Indeed the seventh specimen shows that a mandibular fenestra is absent (Elzanowski & Wellnhofer 1996)
Another important feature of the skull of Archae is the occipital condyle and the foramen magnum. In Archae these are well above the dorsal end of the quadrate. As Ostrom (1976 p. 136) writes:
"This primitive condition is characteristic of both pseudosuchians and theropods, in contrast to all later birds where the occipital condyle and foramen magnum are at the base of the skull, well below the level of the upper extremity of the quadrate. In this feature, Archaeopteryx was far from avian."Whetstone (1983, p. 449) does describe the braincase (as opposed to the skull) of Archaeopteryx as, "typically avian", however, he also describes skull features found in Archaeopyteryx and not in birds.
More recently there has been some conjecture as to the structure of the quadrate bone in Archae. The quadrate from the Eichstatt specimen was described as being single headed - i.e. the top of the quadrate has only one rounded knob which articulates with the skull in only one place. In all modern birds, the quadrate is double headed - i.e. the top of the quadrate has two rounded knobs and therefore articulates with the skull in two places. Based on computer tomography a buried portion of the skull of the Eichstatt specimen, Haubiltz et al. (1988) suggested that Archae possessed a double headed quadrate. However, the image is poor and the presence of another bone underlying the quadrate cannot be ruled out. The only quadrate found in place in any specimen is single headed and a similar bone from the London specimen which appears to be the quadrate, is also single headed. Identification of the quadrate in the seventh specimen has confirmed that the bone was single headed (Elzanowski & Wellnhofer 1996). Thus the quadrate of Archae appears to be reptilian and not avian.
As can be seen, Archae's skull is not "totally birdlike." The "totally birdlike" claim is without foundation.
One is that the differences seen between _Archaeopteryx_, fossil birds and modern birds can be explained as just variation within birds. Allied to this is the claim that some modern birds (Ratites) share a number morphological features with _Archaeopteryx_.
Archaeopteryx + modern birds, just variation within kind?
It has been suggested that the differences between Archaeopteryx and modern birds represents simple within-group variation. However, this is not correct. Modern birds show a large number of derived morphological characters not possessed by Archaeopteryx. Morphologically, Archaeopteryx clearly appears more closely related to theropod dinosaurs that any other group and is grouped with birds over theropod dinosaurs due to the possession of only two main characters, presence of feathers, and presence of a fully reverted hallux (toe).
The relevant morphological variation can be represented graphically (if crudely) in the following figure:
|
^ | A = Archaeopteryx
| |
| ___________________
T | | |
i | | MODERN BIRDS |
m | __ __ __ __ __| |
e | | |___________________|
| | FOSSIL |
| __| BIRDS |
| | A|__ __ __ __ __ __|
| |___|
|______________________________________________
"reptile" "bird"
<--- Morphology -->
As can be seen, the variation within birds shows a distinct trend. The more "reptile"-like morphologies occur in the earliest birds, with the typical 'modern' bird morphology restricted to later birds. If, as suggested, morphological variation is simply variation within birds, we would expect to see the various morphological groups (fossil birds, modern birds) evenly spread throughout the relevant time interval. If, however, birds evolved from theropod dinosaurs, then we would expect to see the first birds to posses more "reptile"-like characters and the more derived, modern birds to have less "reptile"-like characters. This, in fact, is what we see. Thus the distribution of characters within birds supports their derivation from theropod dinosaur ancesters and does not support the claim that variation is simply 'within kind'.
Ratites
Much is made of the morphological features of the Ratites (ostriches, kiwi's etc.), and superficially it may seem that these birds support the 'variation within kind' suggestion as the following figure shows:
_____________________________
| | |
|RATITES| MODERN BIRDS |
|_______| |
|_____________________|
________________________________________________
"reptile" "bird"
<--- Morphology -->
Importantly however, other morphological characters possessed by Ratites clearly show that they are derived from a modern, flying bird morphology and thus the Ratites are actually neotonous on modern birds, with their more "reptile"-like characters due to the early termination of development, leaving them with some morphological characters similar to Archaeopteryx and some fossil birds (see below). These characters are developmental and therefore provide further supporting evidence for a theropod dinosaur-bird link.
A tale of two pelvises
Dinosaurs can be divided into two groups based on the shape of the pelvis. The Saurischia, or 'lizard hipped' dinosaurs, and the Ornithischia, or 'bird hipped' dinosaurs. Strange as it may seem, Archaeopteryx and all modern birds are thought to have evolved from the Saurischia, not from the bird hipped, Ornithischia! Odd? Well, not really. The pelvises in question are shown in Fig.1.
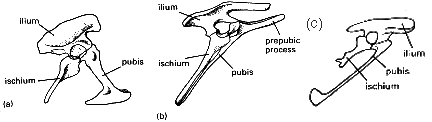
The saurischian pelvis (Fig.1a) is the more primitive structure, and has the pubis pointing forwards and the ischium backwards. In ornithischians (Fig. 1b), the pubis runs back in parallel with the ischium. Additionally, a prepubic process is present, which is assumed to have been used as a point of muscle attachment in the absence of the pubis (probably to support the gut). This situation is superficially similar to that in birds (hence the name 'bird hipped' - the pelvis of birds was well known to anatomists by the time dinosaurs were being described). However, on closer examination, clear differences are evident. In Archaeopteryx (Fig. 1c) and all birds, the prepubic process is absent and, unlike in saurischians, the pubis do not meet along the midline. Also, in Archaeopteryx the pubis still maintains the club-like termination, as in many saurischians. It should also be noted that several saurischian dinosaurs have the pubis facing backward, e.g. Troodon. The pelvis of Archaeopterys is morphologically intermediate between saurischian dinosaurs and birds.
But why did this occur? Well, the primary thrust generator in earlier theropods was the Caudofemoralis longus, which streched from the leg to some distance along the tail. Later forms such as the dromaeosaurids (the group closely related to Archaeopteryx) rearranged this so that the primary thrust was produced by muscles attaching from the leg to the pelvis and the base of the tail. This allowed the tail to be braced and used as a stabilizing device, such as in allosaurs and 'raptors, or to be lost as in modern birds.
In ornithischians the backwards pointing pubis probably occurred to allow for the expansion of the stomach and gut in order to better digest plant material.
Study of the embryonic development of the modern bird pelvis is also instructive. The embryonic pelvis in modern birds starts out in a very similar configuration to Archaeopteryx, with the ilium and pubis bones set apart at an angle of approx. 45 degrees. The embryonic bones then secondarily rotate through angles seen in modern Ratites until reaching the parallel alignment typifing adult, modern birds (see Romanoff 1960, p. 1010):
Chicken enbryo Angle ilium-pubis Adults with same angle
7.0 days 45 degrees Archaeopteryx
7.3 days 35 degrees Kiwi
8.0 days 30 degrees Ostrich
Hatching <5 degrees Modern chicken
The angle between the ilium and the pubis bones in the pelvis of modern Ratites is intermediate between Archaeopteryx and modern birds, as would be expected if:
A) Modern birds are descended from reptile-like ancesters similar to Archaeopteryx
B) Ratites are neotenous descendents of flying birds, in which development has been halted prematurely.
To fly or not to fly
Flying is a tricky business. However, flying confers such a strong evolutionary advantage that it is not surprising that the ability has evolved several times. The success of birds provide ample evidence for the positive benefits of flight. If it did fly, Archae must have utilized much the same method as do birds today, therefore a discussion on the possibility of flight in Archae must consider those structures most relevant to flight in birds. These are the feathers, the flexibility of the wing, muscle bulk and the presence of a keeled sternum.
Feathers
Feathers are composed of a long, tapering, central rachis, which bears closely spaced side branches called barbs. The barbs on either side of the rachis constitute a surface called the vane. The two vanes on the feather may be symmetrical (i.e. the same width) or asymmetrical, in which case the rachis appears closer to one edge of the feather than the other. The flight feathers of modern birds are typically asymmetrical, whilst body contour and semiplume feathers are symmetrical.
In modern birds, remiges, or wing feathers, are highly modified for power flight (e.g. McFarland et al. 1985), primarily in that the rachis is shifted towards the leading edge of the feather (i.e. the leading vane is thinner than the trailing vane), resulting in an asymmetric feather. The thinner or leading vane of the feather overlaps the wider or trailing vane of the feather in front of it (Fig. 2). The trailing vane contains zones of friction barbules which grip the overlapping feather and stop the feathers from slipping too far apart.
Fig. 2
* * * O * * * * *
* * * O * * * * *
* * * O * * * * *
| | |
| | |--trailing vane
| |---------rachis
|-----------leading vane
Speakman & Thomas (1994) compared the asymmetry of some of the flight feathers of Archae (flight feathers 4, 5 and 6) with those of modern flying and flightless birds and the isolated feather from Solnhofen. They found that the average asymmetry for Archae feathers was 1.25, which was lower than that of modern flying birds (lowest around 2.2), but which overlapped that of modern flightless birds. The isolated feather exhibited an asymmetry of 2.2 - just within the range of modern flying birds. However, it is not known whether this feather is from Archae, or where on Archae the feather was situated if it is an Archae feather. Norberg (1995) correctly pointed out, that the curvature of the feather is also important in flight ability, and Archie's feathers exhibit significant curvature. It appears as though Speakmean & Thomas may have measured the wrong feathers however. Paul Davis (pers. comm. 1996) has pointed out that Speakman & Thomas appear to have numbered the primary feathers starting at the wing tip and moving towards the body, whereas current practice is to number the feathers starting from the body and moving outwards along the wing (e.g. Rietschel 1985). This is important, since the asymmetry of feathers decreases towards the wing tip - thus Speakman & Thomas appear to have measured the feathers with lower asymmetry. Measurements of the higher asymmetry feathers close to the boby yield valuse of around 2.32 - which overlap the lower range for extant flying birds (e.g. Norberg 1995; P.G. Davis pers. comm. 1996). The first flight feathers of birds are always highly asymmetric however, and so this result is equivocal. The bulk of the flight feathers on Archae appear to have a slightly lower asymmetry than that found in flying birds today. In reality however, feather asymmetry is a bit of a red herring when it comes to flight capability. Feathers are asymmetrical to maintain aerodynamic integrity as they deform under aerodynamic loads in flight, and thus the greatest asymmetry occurs in birds in which such loads are greatest - i.e. birds with high manouverability and climbing flight (J.M.V. Rayner, pers. comm. 1995). Thus asymmetry is a derived condition for a derived flight capability - one Archaeopteryx never possessed. Thus it is not surprising that Archaeopteryx possessed low asymmetry feathers, but this does not preclude Archaeopteryx from flying.
Wing flexibility
Consider the carniates [birds with a keeled sternum] wing. A marvel of adaptation, well able to sustain quite large animals in power flight for long distances. But what is it about the anatomy of the wing that allows the possibility of flight? Well simply put, 'it's all in the wrist action.'
In modern birds the shoulder joint acts as a universal joint, with a high degree of mobility not only adduction [wing movement down], abduction [wing movement up] and extension [extending wing away from body] and retraction [pulling wing into body], but also significant rotation along the log axis of the humerus.
The elbow, by contrast, is a very restrictive joint. It only allows planar extension and flexure [in other words unfolding and folding of the wing - i.e there is no rotational component.
However, it is the wrist joint which is important. The wrist is composed of two bony carpals which control the movement of the joint. Distally beyond this is the fused carpometacarpus - produced from a fusion of the remaining carpal [wrist] bones and the metacapal [lower hand] bones. Beyond that is the sometimes incompletely fused manus composed of three digits. This complex, distal of the wrist joint, comprises a near-rigid platform for the attachment of the primary remiges [flight feathers], the secondary remiges attach to the forearm.
The configuration of the wrist joint, articulating as it does on two small, round carpels, allows an amazing range of movement. To demonstrate this you need to be sitting comfortably at the computer terminal. Extent your arms as if you were about to type (palms down, hairy bits up). Now, rotate your hands so that the palms are facing up (as if you are carrying something). This rotation is carried out by the wrist joint in birds. Note that in humans, this action is carried out by rotation of the forearm and not the wrist. Now, return the hands to the hairy bits up position and rotate the hands so that the palm faces the computer (the universal "stop" sign) and rotate the hand down until the hairy bits point to the computer. This rotation is carried out by the wrist joint in birds, as it is in humans.
As you have now demonstrated, the carniate wrist is much more flexible than the human wrist (you have also demonstrated to any co-worker watching that you are insane and in dire need of a holiday!). This flexibility is vital to powered flight, since it allows the wrist/hand to describe a lazy figure-of-eight during a complete flap. Remember that the elbow joint allows no rotation, but the wing must be able to present a solid open wing on the down (power) stoke, but be able to rotate the wing during the recovery (up) stroke to minimise resistance. The birds accomplish this by rotating the wrist and shoulder.
At the top of the power stoke, the wrist is orientated to present the maximum surface area to the air (palm facing downward). During the power stroke, the wrist and hand describes an "S", or two arcs representing the left hand side of the upper loop and the right hand side of the lower loop of an "8". At the bottom of the power stroke the wrist and hand is at the bottom of the "8". During the start of the recovery stroke the wrist reorients 90 degrees, thus presenting the minimum surface area to the air. The wing then completes the "8" by describing the left hand side of the lower loop and the right hand side of the upper loop. It is easier to perform than to read. Hold your right arm out horizontally from the shoulder, with the palm pointing downwards. Lock the elbow so that you cannot bend the arm, only rotate it. Now the power stroke is an "S" shape, keeping the palm pointing downwards and the elbow locked. At the bottom of the "S", rotate the palm 90 degrees until it is pointing forwards (actually you will have to rotate your whole arm to do this, in birds it is done by the wrist joint). Now, the recovery stroke is a mirror image "S", completing an "8", as you reach the top of the stroke the palm should twist back to point down again and you are ready for the power stroke. Repeat a few times until you get the feel of it or until someone sends for the guy's in the white coats. That is how birds can fly and, hopefully, you can now see the importance of having a flexible wrist joint.
One last important point in this bit. Folding of the wing occurs by contraction of the M. biceps brachii, which is attached to the ulna. This movement automatically folds the metacarpus and hand downward and inward, so that folding at the wrist joint involves the M. biceps brachii.
Now, the point of all this is: Does this arrangement occur in Archae? Does this arrangement occur in maniraptoran theropods? If so, what possible use could this particular wrist joint be to maniraptoran theropods? Could this be an example of preadaptation (gasp! Sorry to use the "P" word).
Firstly, the wrist joint of Archae does indeed exhibit some (but not all) of the features seen in the modern carniate wrist joint. Most notably a lunate carpal, hemispherical in shape, the flat portion of which articulated with the carpometacarpus, leaving the rounded end to articulate with the wrist joint. This provides a flexible wrist joint, although not as flexible as in modern carniates.
Secondly, this configuration is found in several theropod taxa, such as Deinonychus, Velociraptor and Stenonychosaurus. There are two problems here, firstly all these forms postdate Archae, they are all Cretaceous forms; secondly, what would be the use of a nifty flexible wrist to a bunch of theropods which are all teeth and toenails?
OK, problem one is easy to solve. It was a problem that this feature occurred in Cretaceous theropods and in the Late Jurassic Archae only, since to be an important feature and to show a close link between theropods and Archae, theropods coeval with Archae must have had this particular arrangement. Introducing Coelurus fragilis a late Jurassic theropod from America. This form does indeed possess the a very similar wrist joint (Ostrom 1980) strengthening not only the theropod-bird link, but also the theropod-Archae-bird link.
The second problem is slightly harder. Why would such a wrist joint be retained in maniraptoran theropods which are dentally and toenail minded when it comes to killing? Ostrom (1995) suggests that the flexibility would come in useful where a grasping action of the hands could augment the killing action of the teeth and/or toenails. I would speculate further. In modern birds the biceps are involved in the folding of the wing. If this were extended to the theropods, the action of pulling the prey into the body would tend to strengthen the grip of the theropod by bringing into play the largest muscle on the arm - the biceps, rather than just the wrist muscles. Such a scenario would provide a distinct advantage and thus the retention of the wrist joint in the theropods is not surprising. The title of the sequel to "Jurassic Park" is thus easy: "Jurassic Park II: The cuddle of the Coelusaurs"!
Thus it appears that the flexible wrist joint may have first appeared in theropod dinosaurs as an aid in grasping and retaining prey and was secondarily co-opted by the birds as a mechanism which allowed flight. It may well be that not only were the theropod dinosaurs the ancestors of birds, but that they are the only group which could be the ancestor to modern birds.
However, the wrist of Archae does not appear to have the flexibility to allow power flight.
Shoulder Joint
The shoulder joint is also important to modern birds because, not only does it allow full adduction (wing movement down - Fig. 3b), but also full abduction (wing movement up - Fig. 3d)
Fig. 3 (view head on)
Adduction Abduction
a) -----( )----- c) -----( )-----
b) /( )\ d) | |
| | | |
| | \( )/
i.e. the wing can be drawn up over the back (try it and see how difficult it is). This full range of movement is important in allowing modern birds to power fly. It turns out that the shoulder joint of Archae appears to be intermediate in orientation between some coelurosaurs (theropod dinosaurs e.g. Deinonychus) and birds (Jenkins 1993) - just as one would expect from a transitional form. However, whilst the shoulder joint provided a substantial degree of arm (wing) elevation, it is not sufficient to allow full abduction as in modern birds. Thus the shoulder joint of Archae does not appear to allow the full range of movement needed for power flying.
Muscle mass
In modern birds the flight muscles (the pectoralis) comprise up to 35% of the weight of the animal (think of all that juicy meat at the front end of a roast chicken or turkey). It is this massive muscle mass which provides the power needed for flapping powered flight and the ability to take off from a standing start. Ruben (1991) used a pectoralis muscle mass estimate for Archae of approx. 9% of the body weight. This is much lower than in modern birds, but assumed that the muscle attached to the cartilaginous chest wall and not a keel - since a keel is unknown on Archae. The keel is a bony extension of the sternum for the attachment of excess flight muscle in modern birds. If the muscles did attach to the chest wall in Archae, the length of the pectoralis muscles would have been too short to allow full abduction (see "shoulder joint" above). However, the presence of a small, cartilagenous (and thus not preserved) keel is not out of the question. (Specimens of Archie from the Solnhofen Limestone show no evidence of a sternum. However, recent finds of Archie-like forms from China suggest that Archae did in fact possess a sternum (J.M.V. Rayner, University of Bristol, pers.comm.) Therefore, the 9% figure is a minimum value. This value is well below that for modern birds. Rubin suggested that the maximum power output from the locomotor muscles of extant active reptiles is twice that of birds and mammals, indicating that, if Archae possessed a reptilian physiology, powered flight was a possibility, even with such a small muscle mass. Speakman (1993) questioned this, indicating that the power output of reptiles suggested by Rubin was over-exaggerated, and that Archae would not have had the power output to sustain power flight. Also Speakman pointed out that the theoretical lower limit of pectoralis muscle mass required for take-off from a standing start was approx. 16% of body weight. Thus Archae was not capable of power flight or take-off from a standing start.
The supracoracoideus is an important muscle in modern birds because it is this muscle which elevates or abducts the wing during power flight. Archae did not possess the supracoracoideus arrangement found in modern birds (Ostrom 1974). Thus Archae does not appear to have had the muscle mass nor the muscle arrangement to allow power flight, nor be able to take off from a standing start.
It would appear that the structure and physiology of Archae would mitigate against power flight. But, is this the death knell for Archae's aerobatic abilities? Actually, no. Notice the use of the phrase "power flight" in the preceding discussion. Virtually all modern flying birds power are capable of power flight. Power flight is controlled, flapping flight at low speeds and the ability to take off from a standing start. This type of flying requires a large amount of muscle mass and is energetically very expensive. Hence modern birds have a pectoralis muscle mass which comprises approx. 20-35% of body mass. In order to accommodate this extra mass, a keel has developed on the sternum. In other evolutionary developments, flight feathers have become more asymmetric to assist in power flight, and the supracoracoideus muscle system took on the function of assisting in the lifting or abduction of the wing, since this is difficult at low flying speeds.
Power flight is to flying what the 4 minute mile is to running. However, just as being unable to run a 4 minute mile does not necessarily indicate that one is incapable of running, being unable to power fly does not necessarily indicate that Archae was incapable of flying. The characters concerned with flight we see in modern birds are derived, an adaptation to power flight, and thus their absence (or underdevelopment) in Archae cannot be used to suggest an inability to fly.
The structure of the wing of Archae suggests that it was incapable of power, flapping flight at low speeds. However, the amount of adduction and abduction, along with the increased flexibility of the wrist compared with theropod dinosaurs, suggests that some sort of flapping was possible, enough to keep Archae airborn at normal cruising speeds (approx. 7-9 ms^-1). Whilst the low pectoralis muscle mass appears to preclude power flight and take off from a standing start, it should be pointed out that some flying birds do possess a muscle mass lower that the 16% by weight lower limit. These include some grebes, who compensate for the lack of muscle mass by having a long running take-off (Speakman 1993). Archae is well equipped with a terrestrial locomotion apparatus. Thus it is possible that Archae could circumvent this problem in the same manner as the grebes.
Flying at normal cruising speeds requires much less energy that power, flapping flight and thus needs less muscle mass to sustain it. As an added bonus, at normal cruising speeds the wing is elevated (abducted) in the upstroke by aerodynamic forces (Rayner 1988) - thus circumventing the need for the supracoracoideus arrangement necessary for power, flapping flight.
Archaeopteryx's Ancestry
Apart from the theropod ancestry, there have been three other suggested ancestral groups for Archae, the Ornithopoda, the Pseudosuchia and the Sphenosuchidae.
The Ornithopod ancestry theory was based on the tasus and pelvis in various dinosaurs which were supposed to approach the condition found in birds. However, most of the taxa used in the original theory in 1883-4 are now known to be saurischian rather than ornithischian. Whilst there was a tendency for the reduction of the hallux (big toe) in some ornithischians, it was never apparently opposable as it is in Archae. It was suggested that Archae had a pelvis which was very ornithischian, however, recent work has shown that this is an artifact of post mortem movement of elements of the pelvic region and there is very little similarity to the ornithischian pelvis. With this, the link between ornithischians and Archae disappears and so it is now thought that there is no link between them.
It was suggested that birds arose from an almost unknown Middle or Late Triassic reptile group, the Sphenosuchidae (Walker 1972, 1974), which also gave rise to the crocodiles. The theory is based on a single specimen of Sphenosuchus. However, in their general configurations, neither the skull and jaws, nor the scapulo-coracoid of Sphenosuchus is remotely suggestive of affinities with Archae (Ostrom 1976). More recently, the main proponent of the crocodile-bird hypothesis has stated that the hypothesis "has become so tenuous that it is very difficult to sustain." (Walker 1985, p. 133)
The pseudosuchian link was very much favoured earlier this century. However this was, by and large, based on Heilmann's reconstruction of the Berlin specimen, which has since proven to be inaccurate. According to Ostrom (1976 p. 159):
"There are very few anatomical resemblances between Archaeopteryx and any pseudosuchian. In fact, only in one feature does any pseudosuchian resemble Archaeopteryx more closely than does any theropod, this being the tibia to femur ratio in Scleromochlus, Lagosuchus and Lagerpeton, where the tibia is from 20% to 30% longer than the femur. Amongst theropods, only in struthiomimids, Compsognathus, Microvenator and Deinonychus is the tibia longer than the femur but by only 10% to 15%. In all other features, the closest resemblance to the morphology preserved in Archaeopteryx is found in coelurosaurian theropods." [original emphasis]
Chiappe (1995) provides a very good review of the early evolution of birds.
Protoavis
Some people like to claim that the finding of a fossil bird from the Triassic of Texas (Protavis) proves that Archae cannot be transitional between dinosaurs and birds because Protoavis predates Archae by 75 million years. This is, of course, errant nonsense, mainly because no one is claiming that Archae is the transitional species between dinosaurs and birds, merely that Archae represents a grade of organisation which the proposed lineage went through to get from dinosaurs to birds. Archae is, I'm sorry to say, out on a limb, evolutionarily speaking. It represents a side branch, useful for comparative purposes, but not in the thick of things. So even if there were birds in the Triassic, that fact would not diminish Archae's importance as an indicator that "yes, birds could have evolved from dinosaurs."
However, notice the "if" in the previous sentence. There are major problems with Protoavis. On the Chatterjee (1991) interpretation, Ostrom (1991) has this to say [my paraphrase]: The only published material from the fossil is a monograph in the Philosophical Transactions of the Royal Society of London. However, this only describes the head. This is badly crushed and all the pieces have been extracted from the matrix, rendering precise placement of the pieces open to question. The description is done from an avian viewpoint, with no counterview (e.g. is this a dinosaur?) used. The skull is so badly crushed that diagnostic features are not preserved. Therefore the published material does not support the view that this is a bird. Indeed a viewing of the fossil by Ostrom (in admittedly less than ideal surroundings) showed that the diagnostic features which could identify the fossil either way are badly crushed and it is doubtful whether any definitive statement could be supported by the fossil. It may be a bird, it may not.
Please note that this questioning of Protoavis as a bird is no "it can't be a bird because it predates Archae" evilutionist backlash. As has been pointed out, even if it is a bird, it does not detract from the evolutionary importance of Archae.
Conclusions
Archaeopteryx is a bird because it had feathers. However, it retained many dinosaurian characters which are not found in modern birds, whilst having certain characters found in birds but not in dinosaurs. By virtue of this fact Archaeopteryx represents an example of a group in transition - a representative which, although on the sidelines in the dinosaur to bird transition, an echo of the actual event, still allows a brief glimpse into the possible mechanism which brought about the evolution of the birds and by its very existence shows that such a transition is possible.
Acknowledgements
This came about as the result of a series of discussions with Rich Trott, who was fighting the systematic misuse of Archae by certain opponents of evolution, and still managed to find the time to suggest many improvements to this post. The contributions of Tom Holtz and Paul Davis are also acknowledged. Most of the original version was coverted to HTML format for the talk.origins archive by Brett Vickers.
This is a University of Ediacara Palaeontological Contribution.
References
Abbott, A. (1992) Archaeopteryx fossil disappears from private collection. Nature, 357: 6.
Alexander, R.M. (1990) Animals. Cambridge University Press, Cambridge. 509pp.
Barsbold, R. et al. (1990) Oviraptorosauria. In The Dinosauria, Weishampel, Dodson &p; Osmolska (eds) pp 249-258.
Barthel, K.W.; Swinburne, N.H.M. & Conway Morris, S. (1990) Solnhofen. A study in Mesozoic palaeontology. Cambridge University Press, Cambridge. 236pp.
Bellairs, A.D'A. & Jenkin, C.R. (1960) The skeleton of birds. p. 241-300. In: Biology and Comparative Physiology of Birds, Vol. I, A.J. Marshal (ed.). Academic Press, New York.
Britt, B.B.; Makovicky, P.J.; Gauthier, J. & Bonde, N. (1998) Postcranial pneumatization in Archaeopteryx. Nature, 395: 374-376.
Brooks, B. (1993) Pneumatic postcranial bones in dinosaurs and other archosaurs. Ph.D. Dissertation, Department of Geology and Geophysics University of Calgary, Calgary, Alberta, Canada.
Bryant, H.N. & Russell, A.P. (1993) The occurrence of clavicles within dinosauria: implications for the homology of the avian furcula and the utility of negative evidence. Journal of Vertebrate Paleontology, 13(2):171-184.
Chatterjee, S. (1991) Cranial anatomy and relationships of a new Triassic bird from Texas. Phil. Trans. R. Soc. Lond. B. 332: 277-342.
Chen Pei-Ji; Dong Zhi-Ming & Zhen Shuo-nan. (1998) An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature, 391: 147-152.
Chiappe, L.M. (1995) The first 85 million years of avian evolution. Nature, 378: 349-355.
Chure, D.J. & Madsen, J.H. (1996) On the presence of furculae in some non-maniraptoran theropods. Journal of Vertebrate Paleontology, 16: 573-577. Colbert, E. H. & Morales, M. (1991) Evolution of the vertebrates : a history of the backboned animals through time. 4th ed. Wiley-Liss, New York. 470 p.
de Beer, G. A. (1954) Archaeopteryx lithographica. British Museum of Natural History, London.
de Beer, G.A. (1956) Evolution of the Ratites. Bulletin of the British Museum (Natural History): Zoology, 4: 57-70.
Elzanowski, A. & Wellnhofer, P. (1996) Cranial morphology of Archaeopteryx: Evidence from the seventh skeleton. Journal of Vertebrate Paleontology 16 (1): 81-94
Gingerich, P.D. (1975) Evolutionary significance of the mesozoic toothed birds. Smithsonian Contributions to Paleobiology, 27: 23-34.
Haubiltz, B.; Prokop, M.; Dohring, W.; Ostrom, J.H. & Wellnhofer, P. (1988) Computed tomography of Archaeopteryx. Paleobiology, 14: 206- 213.
Heilmann, G. (1926) The Origin of Birds. Witherby, London. 208pp.
Jenkins, F.A. (1993) The evolution of the avain shoulder joint. American Journal of Science, 293A: 253-267.
Lambrecht, K. (1933) Handbuch der Palaeornithologie. Berlin. 1024 pp.
McFarland, W.N.; Pough, H.F.; Cade, H.F & Heiser, J.B. (1985) Vertebrate Life, 2nd ed. Macmillan Publishing, New York. 636 pp.
McGowan, C. (1984) In the Beginning: A Scientist Shows Why the Creationists are Wrong. Prometheus, Buffalo. 208 pp.
Norberg, R.A. (1995) Feather asymmetry in Archaeopteryx. Nature 374: 221.
Orr, R.T. (1966) Vertebrate Biology, 2nd ed. W.B. Saunders, Philadelphia. 483pp.
Ostrom, J.H. (1970) Archaeopteryx: Notice of a "new" specimen. Science, 170: 537-538.
Ostrom, J.H. (1972) Description of the Archaeopteryx specimen in the Teyler Museum, Haarlen. Proceedings Koninklijke Nederlandse Akademie Van Wetenschappen, B, 75: 289-305.
Ostrom, J.H. (1974) Archaeopteryx and the Origin of Flight. The Quarterly Review of Biology, 49(1)
Ostrom, J.H. (1976) Archaeopteryx and the origin of birds. Biological Journal of the Linnean society, 8(2): 91-182.
Ostrom, J.H. (1980) Coelurus and Ornitholestes: Are they the same? In L. Jacobs (ed) Aspects of Vertebrate History. 245-256.
Ostrom, J.H. (1991) Bird in the Bush. Nature, 353: 212
Ostrom, J.H. (1995) Wing biometrics and the origin of bird flight. Neues Jahrbuch fur Geologie und Palaeontologie, 195: 253-266.
Padian, K. (1998) When is a bird is not a bird? Nature, 393: 729-730.
Qiang Ji; Currie, P.J.; Norell, M.A. & Shu-an ji. (1998) Two feathered dinosaurs from northeastern China. Nature, 393: 753-761.
Rayner, J.M.V. (1988) The evolution of vertebrate flight. Biological Journal of the Linnean Society, 34: 269-287
Rietschel, S. (1985) In: The Beginnings of Birds, Hecht, M.K., Ostrom, J.H., Viohl, G. & Wellnhofer, P. (eds.). pp. 303-318. Freunde Des Jura-Museums, Eichstatt.
Romanoff, A.L. (1960) The Avian Embryo. Macmillan, New York. 1304pp.
Romer, A.S. (1950) Vertebrate Paleontology. p 261. University of Chicago Press, Chicago.
Ruben, J. (1991) Reptilian physiology and the flight capacity of Archaeopteryx. Evolution, 45: 1-17.
Speakman, J. R. (1993) Flight capabilities in Archaeopteryx. Evolution, 47: 336-340.
Speakman, J.R. & Thomson, S.C. (1994) Flight capabilities of Achaeopteryx. Nature, 370:514.
Tucker, B.W. (1938a) Functional evolutionary morphology: the origin of birds. p. 321-336. In: Evolution: Essays on Aspects of Evolutionary Biology, G.A. de Beer (ed.). Clarandon Press, Oxford.
Tucker, B.W. (1938b) Some observations on Dr. Lowe's theory of the relationship of the Struthoines to the dinosaurs and to other birds. Proceedings of the 8th International Ornithological Congress, Oxford. 1934: 222-224.
Unwin, D M (1998) Feathers, filaments, and theropod dinosaurs, Nature 391: 119-120.
Walker, A. D. (1972) New light on the origins of birds and crocodiles. Nature, 237: 257-263.
Walker, A. D. (1974) Evolution, organic. In Yearbook of Science and Technology, D.N. Lapedes (ed). pp. 177-179. McGraw-Hill, New York.
Walker, A. D. (1985) The braincase of Archaeopteryx. In The Beginnings of Birds, Hecht, M.K., Ostrom, J.H., Viohl, G. & Wellnhofer, P. (eds.). pp. 123-134. Freunde Des Jura-Museums, Eichstatt.
Wellnhofer, P. (1974) Das funfte skelettexemplar von Archaeopteryx. Palaeontographica A, 147: 169-216.
Wellnhofer, P. (1993) The seventh specimen of Archaeopteryx from the Solnhofen Limestone. Archaeopteryx, 11: 1-47.
Whetstone, K. N. (1983) Braincase of Mesozoic birds: I. New preparation of the "London" Archaeopteryx. Journal of Vertebrate Paleontology, 2: 439-452.
Witmer, L. M. (1990) The craniofacial air sac system of Mesozoic birds (Aves). Zoological Journal of the Linnean Society, 100: 327-378.