Summary: Biology cannot be reduced to physics, even though all biological entities are physical entities, and nothing more. Group selection is not an accepted evolutionary theory, but group sorting is.
Previous |
Contents |
Next |
![]() he philosophy of science, and
criticisms of evolutionary theory in particular, has been
driven by the view that physics, or perhaps mathematics, is
the very model of a modern scientific discipline. If it
ain't like physics in some way, then it ain't science.
he philosophy of science, and
criticisms of evolutionary theory in particular, has been
driven by the view that physics, or perhaps mathematics, is
the very model of a modern scientific discipline. If it
ain't like physics in some way, then it ain't science.
Unsurprisingly, many biologists weren't happy with this view of what they do as some kind of 'stamp collecting' [note 7]. Ernst Mayr [1970, 1982] attacked this philosophical presumption, especially the idea that biology is just a form of physics, or perhaps chemistry. Philosophers, too, started to make similar attacks [Hull 1974, see Sterelny 1995 for a review].
The view of science of philosophy in the early 60s was generally reductionist [Nagel 1961]. This meant that in principle the objects and processes of one level of science were made out of the objects of the next level down, ending with subatomic physics. Thus, biology reduced to chemistry, and chemistry reduced to physics. This sort of reduction is called ontological reduction. Those who accept this form of reductionism are called 'physicalists'.
Another type of reduction - often confused with the ontological - is explanatory or epistemic reduction. This is the view that the properties of one level must be ideally explained as the effects of processes at the next level down. This is roundly denied by many philosophers and biologists, and asserted by many others [eg, Dennett 1995]. Hull [1974] argued that it is in principle impossible to reduce, for instance, population genetics to Mendelian genetics, and Mendelian genetics to molecular genetics, because each level is the result of many entities interacting at the lower level, and many entities at the higher level result from a single entity at the lower level:
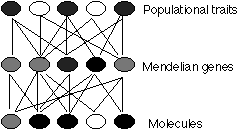
Hull's problem is sometimes called the Problem of the Many to Many, in homage to an old philosophical problem, the Problem of the One and the Many. Many Mendelian genes are made from many DNA molecules, and many populational traits are coded for by many Mendelian genes. Simple reduction will not work. What Williams [1966] called an 'evolutionary gene' is just a unit of heredity that is 'visible' to selection, and it could be an entity at any level - a molecule, a Mendelian gene or even a populational trait.
Reduction enters the evolutionary debate in the form of the issue of group selection. In 1962, Wynne-Edwards proposed that some bird populations regulate their clutch size (the number of eggs laid) in hard times to benefit the population as a whole, even though it was detrimental to the 'Darwinian fitness' of the individual birds. Williams [1966] responded with an argument that selection of individuals could not account for this and other forms of supposed group selection, and that if group selection occurred at all, it was not very important.[note 8] A decade later, Dawkins [1976] hardened this view into the claim that genes, and genes alone, are the 'units of selection', and that all biological effects in evolution are the result of these 'lower-level' entities.
Gene-centrism is not the view that only genes exist, or even that only genes have effects, but that only genes are selected (that is, are evolutionarily important). The way Dawkins put it, as evidenced by the title The Selfish Gene, was wrongly interpreted to mean that organisms are irrelevant. More informed analysis developed the view that if evolution-by-selection is generalised, then using Dawkins's own distinction between replicators and vehicles (or Hull's refinement, interactors), then selection can occur at levels above the gene, or even above the organism.
This puts the lie to simplistic notions that evolution is defined as a change in allele frequencies. That is what has been called by Wimsatt the 'bookkeeping' definition of evolution, and it is true as far as it goes, but it is not all that is interesting about evolution. Few biologists are still simple reductionists, although Williams did write a limited defence of reductionism as a methodological ploy [1985], in which he argued that reduction was the 'null hypothesis' and it was onerous to abandon it. Dennett [1995] claimed that reduction had yet to fail, especially in evolutionary explanations using selection.
If evolutionary levels above the gene can be selected, are they adapted? Several have followed Wynne-Edwards on this. Recent versions, though, have moved from the notion that groups are selected, though, in favour of the view that they are sorted, because selection requires that the entity in question replicates, and does so differentially relative to other contenders. This is OK for genes, and arguably for organisms, but species? Even phyla? Groups do not reproduce, they split. Recent work by Gould, Eldredge and Vrba [refs in Sterelny 1995] amends this from species selection to species sorting, what Vrba calls the 'effect hypothesis'. This is a view that now has wide acceptance amongst biologists, including Williams [1992]. Groups are thought to survive extinction events differentially, based on adaptations of their component organisms, so the organisms are adapted, not the groups.
Previous |
Contents |
Next |

Home Page | Browse | Search | Feedback | Links
The FAQ | Must-Read Files | Index | Creationism | Evolution | Age of the Earth |
Flood Geology | Catastrophism |
Debates
