
Response to Casey Luskin
by Nic Tamzek et al.Copyright © 2002
[First Posted: March 10, 2002]
[Last Updated: March 14, 2002]
[Last Updated: March 14, 2002]
![]()
![]()
Other Links:
|
![]() he first incarnation of Nick
Tamzek's essay "Icon of
Obfuscation" was a rather long flier written to be
passed out at a lecture by
Jonathan Wells at UCSD on January 29th, 2002. The text of
this flier was posted on the web shortly thereafter, and
several versions of additions and edits were posted in
succession until the essay was posted on talkorigins.org on
February 25th, 2002.
he first incarnation of Nick
Tamzek's essay "Icon of
Obfuscation" was a rather long flier written to be
passed out at a lecture by
Jonathan Wells at UCSD on January 29th, 2002. The text of
this flier was posted on the web shortly thereafter, and
several versions of additions and edits were posted in
succession until the essay was posted on talkorigins.org on
February 25th, 2002.
Shortly afterwards, Casey Luskin of the UCSD IDEA club, one of the organizers for Dr.Wells' lecture at UCSD, posted a response to the original flyer (which he is hosting here) entitled "Icons Still Standing: Jonathan Wells Comes Up Clean, Despite Harsh Criticism". The same essay has thus far been also been posted at the San Diego IDEA center website and at the ID site Access Research Network (ARN).
While the original flier lacked much of the material that is in the Talk Origins FAQ, Luskin's lengthy response does attempt to address many key arguments of the current FAQ, and so far it is Wells' only known defense. So a response to Luskin is worthwhile. All the same, Dr. Wells is urged to respond for himself.
This response has been assembled by the somewhat novel technique of having numerous writers respond to specific pieces of Luskin's essay. One reason for this is to post a version of it by March 11, when Wells and others speak in Ohio. Luskin has offered such a long laundry list of creationist claims that one person cannot straighten them all out in a short time, even though they are not new. This may make it a bit disjointed to read, but it allows for various people to contribute in areas of relevant knowledge.
In order to keep things clear, quotes of Luskin will be in gray boxes like this. All quotes are copied directly from the version of Luskin's essay that was saved from the web on March 3rd, 2002. A few typos in the original will therefore be left as is, without [sic]s, etc. The version of Luskin's defense of Wells that is posted at the ARN site and the IDEA site keeps changing as people point out errors, so this response of necessity refers to the March 3rd version. Furthermore, note that this document is a work in progress. Some of what Luskin wrote will be addressed in future versions of this document.
Abelson goes on to say that there is "no evidence for ... but much against" an ammonia-methane atmosphere, noting that, "[a] quantity of ammonia equivalent to present atmospheric nitrogen would be destroyed [by ultraviolet radiation] in ~30,000 years."2 He, and Lasaga et. al. found that methane gas would have left a signature of organic carbon in ancient sedimentary rocks, perhaps even due to a 1 to 10 m thick "primordial oil slick"15 (after all, Miller-Urey experiments produced by far more tar than anything else, and if many prebiotics were produced, then tar should have covered the early earth), for which there is no geological evidence. Schopf notes that the Swaziland Supergroup (~3.2 Ga) contains inorganic carbon, "indicating that carbon dioxide, and not methane, may have been the dominant form of atmospheric carbon at this point in time"16. This challenges the importance of undersea thermal vents as a contributor of methane, which Tamzek says Wells "exclude[s]" wrongly.
As this is around 300 million years after the first known bacterial fossils, and well after the time frame of the origin of life (4.2-3.8 Bya), is anyone surprised. Furthermore, in Kastings model, methane is a minor (around 20 ppm) not dominant form in the atmosphere, so this is doubly irrelevant to the hydrothermal vent model.
-- Ian Musgrave
Darwin's Clock is Ticking
Tamzek notes that the molecular clock hypothesis is in great dispute, which is most pronounced when the molecular clock differs markedly from the fossil record (especially with regards to mammals36, birds36, and major animal groups36, 37). Cone snails79, which Tamzek allegedly cites as an example of rapid evolution, provide little fodder for refuting creationist claims against the natural origin of genetic information, as it merely shows that there was strong selection pressure on cone snail mrRna. If evolution did occur, no novel functions were created apart from a few different destructive venoms--and these venoms have themselves had between 20-50 Ma to develop into their fairly similar forms. The paper actually admits its most striking example of alleged rapid change, which it calls a "focal hypermutation,"79 is "mechanistically unexplained", making naturalistic explanations, at this point, impotent to account for the alleged genetic transformations.
When Luskin gets around to addressing gene duplication as an example of evolution increasing information, he has some bizarre things to say.
This paper, like many others in the literature of evolutionary biology, bases most of its genetic evolution of biochemistry off of the mechanism of gene duplication. However, when trying to evolve something, the gene-duplication-explanation doesn't help the issue much, because once you duplicate a gene, you have a new piece of genetic information to play around with, but what good is that to you? If complex systems need specific parts, what sort of evidence is there that these duplicated genes will be the parts you need?
Of course, gene duplications occur in already living organisms that don't "need" anything. However, if a duplication produces an additional useful function, or expands upon an existing function, then natural selection will help it spread. This is one of the more frequent ways that new functional information gets added to genomes, and new complex systems are the eventual outcome of new genes working together.
Luskin bases most of his dismissal on a paper by Lynch and Conery [1], claiming that the rate of gene duplication is too small, but his use of the paper has several serious problems:
1. Lynch and Conery, by using whole genomes, have tried to empirically derive the rate of single gene duplications, and have done so excluding multi-gene families. Their study did not address whole genome duplication, partial or whole chromosomal duplication, or other kinds of segmental duplication. Luskin dismisses duplication as a means for generating biological novelty, but he fails to address the more potent mechanisms for the addition of genetic material, which are capable of adding hundreds or thousands of genes at a time.
2. Contrary to Luskin's contention that the rate of duplication reported is "pretty rare," it is actually considered quite high. Lynch and Conery state that "50% of all of the genes in a genome are expected to duplicate and increase to high frequency at least once on time scales of 35 to 350 million years. Thus, even in the absence of direct amplification of entire genomes ( polyploidization), gene duplication has the potential to generate substantial molecular substrate for the origin of evolutionary novelties."
3. Lynch and Conery's methodologies and conclusions were both heavily criticized in a Science "technical comments" piece by two separate sets of scientists [2], [3] who are well regarded experts. Among other things, the conclusion of the short half-life of gene duplicates and the rate of duplication were both questioned, and while Lynch and Conery defend their position, they admit that several assumptions bias their results and they revise some of their estimates. So the results of the study are at the very least highly questionable. Luskin should have either mentioned this controversy, or left out the study all together.
Even without these problems, Luskin still fails to adequately address the issue of duplication by relying on this study. The rate that Lynch and Conery present, even if correct, is an average rate of duplication. But various genes can deviate greatly from the norm. Furthermore, it is well known that rates of gene duplication are not uniform. For example, subsequent duplications are much more likely if a duplication has occurred previously, because the chances of unequal crossing over are greatly increased [4], This allows for the rapid expansion of gene families, which appears to be what happened in the case of the cone snail venoms. Lynch and Conery left gene families out of their study.
Lynch and Conery found that the average gene duplicates about once every 100 million years108--that's pretty rare. If cone snails have a 1 year generation time, and the gene you need duplicates once every 100,000,000 generations, that doesn't give you a very good chance of getting it when you need it. Furthermore, it has been found that, "the vast majority of gene duplicates are silenced within a few million years, with the few survivors subsequently experiencing strong purifying selection"108. Another study showed are not very free to mutate around at all, that there is strong selection pressure on them107. This supports the statement by Conery and Lynch that the actual mechanisms by which gene duplication contributes to evolution are not very well understood:
"However, it is unclear how duplicate genes successfully navigate an evolutionary trajectory from an initial state of complete redundancy, wherein one copy is likely to be expendable, to a stable situation in which both copies are maintained by natural selection. Nor is it clear how often these events occur."108
Luskin draws on Lynch and Conery to claim that new duplicates don't last long because they're quickly (i.e. millions of years) silenced by mutation. (Ironically, the ones that get silenced become pseudogenes, which offer powerful evidence of common descent. See "Plagiarized Errors and Molecular Genetics", http://www.talkorigins.org/faqs/molgen/). But aside from the aforementioned problems with that study, Luskin simply ignores recent studies that show that duplicates are preserved far more often than previously thought [5], [6] including studies by Lynch himself [7], [8]. In addition to evolving completely new functions, gene duplications can partition old functions that the ancestral gene carried out only weakly. This model is supported by the fact that many enzymes are "promiscuous" in their number of weak activities [9]. This allows the partitioned functions to be more active and differently regulated, sometimes being expressed in different tissues [10]. In this way, greater complexity can evolve even without de novo functions being added, and duplicates are more frequently preserved. Luskin concludes:
The bottom line is that the gene duplication explanation still leaves the details to the dice, and this pathway definitely hasn't been experimentally verified. All Espiritu et al. have found are protein homologies, and then inferred a vague ancestral pathway of gene creation. This explanation for the origin of real evolutionary novelty lacks a reliable mechanism and is little better than hand waving.
But it is clear that it is Luskin who is doing the hand waving. He relies on a questionable study that he misinterprets while ignoring a wealth of data that contradict his thesis. Contrary to his claims, the mechanisms for gene duplication, which include unequal crossing over and the insertion of retrogenes, are reasonably well understood [4] and are followed by any number of selective possibilities (see any of the above or below references for extensive information, or any of dozens of others). Furthermore, adaptive gene duplication has been witnessed in the laboratory [11], [12]. And observed between closely related species [13] (including cone snails [14]) that even young-earth creationists consider related by common descent. Certainly, there are more things to be learned about gene duplication (which is one reason why cone snail venoms are so heavily studied), but whether or not it occurs, or its importance in evolution, is not in doubt. It is simply wrong to claim that gene duplication is not a reliable mechanism for generating biological novelty.
REFERENCES
1. Lynch, M. and J.S. Conery, The evolutionary fate and consequences of duplicate genes. Science, 2000. 290(5494): p. 1151-5.
2. Long, M. and K. Thornton, Gene duplication and evolution. Science, 2001. 293(5535): p. 1551.
3. Zhang, L., B.S. Gaut, and T.J. Vision, Gene duplication and evolution. Science, 2001. 293(5535): p. 1551.
4. Patthy, L., Protein Evolution. 1 ed. 1999: Blackwell Science Inc. 91-103.
5. Massingham, T., L.J. Davies, and P. Lio, Analysing gene function after duplication. Bioessays, 2001. 23(10): p. 873-6.
6. Dermitzakis, E.T. and A.G. Clark, Differential selection after duplication in mammalian developmental genes. Mol Biol Evol, 2001. 18(4): p. 557-62.
7. Force, A., et al., Preservation of duplicate genes by complementary, degenerative mutations. Genetics, 1999. 151(4): p. 1531- 45.
8. Lynch, M. and A. Force, The probability of duplicate gene preservation by subfunctionalization. Genetics, 2000. 154(1): p. 459- 73.
9. James, L.C. and D.S. Tawfik, Catalytic and binding poly- reactivities shared by two unrelated proteins: The potential role of promiscuity in enzyme evolution. Protein Sci, 2001. 10(12): p. 2600-7.
10. Serluca, F.C., et al., Partitioning of tissue expression accompanies multiple duplications of the Na+/K+ ATPase alpha subunit gene. Genome Res, 2001. 11(10): p. 1625-31.
11. Jelesko, J.G., et al., Rare germinal unequal crossing-over leading to recombinant gene formation and gene duplication in Arabidopsis thaliana. Proc Natl Acad Sci U S A, 1999. 96(18): p. 10302-7.
12. Brown, C.J., K.M. Todd, and R.F. Rosenzweig, Multiple duplications of yeast hexose transport genes in response to selection in a glucose-limited environment. Mol Biol Evol, 1998. 15(8): p. 931-42.
13. Cirera, S. and M. Aguade, Molecular evolution of a duplication: the sex-peptide (Acp70A) gene region of Drosophila subobscura and Drosophila madeirensis. Mol Biol Evol, 1998. 15(8): p. 988-96.
14. Espiritu, D.J., et al., Venomous cone snails: molecular phylogeny and the generation of toxin diversity. Toxicon, 2001. 39(12): p. 1899-916.
-- Steve Reuland
Save the trees!
Tamzek implies that Darwin's tree of life looks simple and "tree-like" apart from the 'molecular thicket' at the root, however this is not the case. It is well recognized in systematics that very often phylogenetic trees based upon one gene or protein sequence, will lead to one tree, while a tree based upon some other biomolecule look quite different. And even more often, trees based upon biomolecules are at odds with trees created on macromorphological characteristics or the fossil record (and trees based upon different macromorphologies often differ from one another a well). Tamzek claims that the Eukaryote tree is well resolved through Baldauf et al. (2000)99, however this paper escapes typical problems of conflicting trees because it creates one tree using a massive dataset of many protein sequences to statistically obscure the differences between the trees based upon individual proteins. By creating one "flimsy" tree through many genes, common ancestry cannot be strongly verified through independent converging lines of genetic evidence (admittedly, this is a good technique for creating a phylogeny if one already assumes common ancestry is true, however, as a test of common ancestry, it shows molecular evidence provides little support). In fact, it was found that single-gene phylogenies only "support subsets of the combined protein tree." In other words, the different genes, when taken independently, do not converge to make a nice neat tree.
Luskin's claim that Baldauf et al. combined the data to obscure conflict is baseless. The authors explicitly recognized the potential for conflict between data sets, and in order to address this issue they analyzed the proteins separately before combining them. Here is what Baldauf et al. wrote:"
Analyses of individual proteins show that no conflicting groupings are supported by >65% aaBP and most by much less (Fig. 3, columns 1 through 4). Therefore, no strongly supported conflicts exist among these data. Pairwise analyses also show considerable evidence of phylogenetic cooperation among all partitions for most questions (Fig. 3, columns 5 through 10); i.e., most pairwise combinations tend to support the same conclusions. This is further evidence of common underlying histories among all partitions. [Emphasis added.]
Baldauf et al. didn't combine the data to obscure conflict because they didn't find significant conflict. The clades that appeared to conflict did not have statistical support, and were probably the result of insufficient data.
- Alex Wild
Tamzek claims that the results of this article indicate that Eukaryote phylogeny is coming along "just fine", and that this paper provides a "perfectly traditional and tree-like" phylogeny of the Eukaryotes. As said before, the neatness of this tree is purely the result of statistical techniques and the combined dataset used in the paper.
Looks like we must add statistics to the list of disciplines that must be flawed if I.D. arguments are true. The primary "statistical technique" used to produce "neatness" of the Baldauf et al. tree is the use of a larger sample of character data. If drawing from a larger sample is an underhanded statistical technique, pigs fly.
Following Luskin's logic, we ought to reject the result of a 100 person opinion poll in favor of the result of a 10 person opinion poll, because the former used a "statistical technique" that produced a different result. Luskin would have us weigh a weakly supported result based on a small sample equally to a much more robust study based on a larger sample.
Why does he do this? I don't know. I suppose sampling error produces enough strange results that I.D. proponents can find some to use for rhetorical purpose, especially if they sidestep the issues of sample size and statistical power. However, once we begin weighing studies not on the merits of their methodology but by how well we can use them in our rhetoric we cease being good scholars and start being propagandists.
-- Alex Wild
Regardless, Baldauf et al. actually found that their tree has "striking differences from SSU rRNA phylogeny"99, and noted that past studies have also encountered conflicting trees.
Let's look at the abstract of the Baldauf et al. paper, the source of the quote:
Current understanding of the higher order systematics of eukaryotes relies largely on analyses of the small ribosomal subunit RNA (SSU rRNA). Independent testing of these results is still limited. We have combined the sequences of four of the most broadly taxonomically sampled proteins available to create a roughly parallel data set to that of SSU rRNA. The resulting phylogenetic tree shows a number of striking differences from SSU rRNA phylogeny, including strong support for most major groups and several major supergroups. [Emphasis added.]
Notice in particular the phrase that Luskin omitted:
...including strong support for most major groups and several major supergroups
The ommitted text changes the meaning of the sentence significantly. Luskin was making the case that the SSU rRNA gives a different tree than that produced by the 4 proteins used by Baldauf et al., and in particular was arguing that the two data sets produce conflicting trees. However, in context it is clear that the differences between the SSU rRNA and the Baldauf et al. trees are not primarily in branching order, but in level of support for the major groupings. Further reading of Baldauf et al. bears this out. The difference between strong support and weak support for a grouping (or of strong support for a grouping versus weak support for a different grouping) is most assuredly not the same as the difference between two strongly-supported but conflicting groupings. Luskin's argument rests on conflating the two.
Source: Baldauf SL, Roger AJ, Wenk-Siefert I, Doolittle WF. "A kingdom-level phylogeny of eukaryotes based on combined protein data." Science 290(5493):972-7 (2000 Nov 3).
-- Alex Wild
Developmental biologist PZ Myers was asked via email to comment on what Luskin wrote:
The Embryonic Hourglass Figure
Tamzek graciously concedes that Haeckel's drawings do not belong in textbooks (essentially affirming the icon), however, he cites Raff (2001)30 to imply that embryology still provides evidence for a shared evolutionary history among vertebrate classes. Raff implies Wells denies the "phylotipic stage" and "sharing of major structural elements and their topological elements" among vertebrate embryos, however Wells readily acknowledges that embryo similarities of the "phylotipic stage" do exist saying, "here [at the phylotipic stage] the various classes first exhibit the characteristics common to all vertebrates" (Icons, pg. 98). Wells' point, here, is that the paths developing vertebrates take to get to the "phylotipic stage" are very different, forming a pattern of development Raff himself says is shaped like an "hourglass"32. Wells notes this "hourglass" pattern is not predicted if all vertebrates share a common ancestor.
This is correct. A specific pattern of development is not an evolutionary prediction; the developmental hourglass is an observation. We see a phylotypic period over and over again in different species, which raises an interesting question: why do animals do that? The evolutionary explanation is that there is a morphological foundation (the body plan) that is set up at an early period in development, which has been refractory to change. Later elaborations on the body plan are based on the patterns laid down in the phylotypic stage, which is why that stage, broadly defined, persists.
Furthermore, we can make predictions based on hypotheses about how body plans are set up. We know that Hox genes are central to this process. One prediction that can be made is that organisms with different body plans ought to have correlated changes in the way their Hox genes are expressed, or in the way Hox genes regulate genes further downstream. These predictions have been tested and are currently being tested in labs around the world.
The creationist/ID camp lacks any explanation for the common features we see in widely disparate organisms. If their 'theories' had any power, they should be able to help us understand these observations. They don't.
Myers:
[...]
Richardson says this is not a problem for Darwinian evolution because "the mixture of similarities and differences among vertebrate embryos reflects evolutionary change in developmental mechanisms inherited from a common ancestor". However, looking at the data Wells presents in his Homology in Vertebrate Limbs chapter, we find that mechanisms controlling development are often not at all predicted by the phylogeny. Wells finds that the gene Distal-less controls development of appendages in the mouse, spiny worm, butterfly, sea urchin, and velvet worm, even though such appendages are not derived through a common ancestry. There is no reason why common ancestry would predict that the same gene would produce "legs" on so many very different types of organisms, when their alleged last common ancestor supposedly did not have legs.
This is partially true. It is correct that we would not predict that legs in arthropods and vertebrates would necessarily have the same molecular cause, because legs in those two lineages are not homologous -- they arose independently. However, what we have here again is an observation. Vertebrate and arthropod legs all use the same gene, distal-less, to initiate some of the early steps in limb development! That is a rather surprising result that favors an evolutionary explanation. Evolutionists have always predicted some unspecified level of similarity between different lineages, because they arose from a common ancestor, but we couldn't know how similar they would be until we looked. Now we're in the curious position creationists are trying to argue that because the similarities between organisms are stronger than we had anticipated, that is evidence against the idea of common descent!
Vertebrate paleontologist Thomas R. Holtz, Jr. who showed just how inaccurate Luskin is. Holtz wrote in an email:
Grounded.
Tamzek claims that fossil theropod Dromaeosaurs (Protoarchaeopteryx and Caudipteryx) provide unequivocal evidence that birds evolved from dinosaurs.
Neither Protarchaeopteryx (note spelling) nor Caudipteryx are dromaeosaurs. They are a generalized maniraptoran and an oviraptorosaur, respectively.
In the remainder of this section the replies to Luskin in quotes are from the email from Holtz. He provided the following cladogram:

"On this diagram, the following groups have confirmed
feathers/feather analogs:
Avialae (birds) (duh...)
Dromaeosauridae (Microraptor,
Sinornithosaurus, and the new form mentioned in
Nature this week)
Alvarezsauridae (Shuvuuia)
Oviraptorosauria (Caudipteryx)
Therizinosauroidea (Beipiaosaurus)
Compsoganthidae (Sinosauropteryx)
Note that other than Shuvuuia and birds, all the
discoveries are from a single Formation, the Yixan, noted
for its spectacular preservation. Note also that we do not
have scale impressions or any non-feather impressions for
other species in these particular groups, so the simplest
explanation is that their most recent common ancestor had
simple feathers."
"The dinosaurs from Tyrannosauroidea up to birds on this cladogram are noted by a reduction in the number of tail vertebrae; this is particularly true of birds and their closest relatives."
However, the skeptic must ask the question, is this really the case? These are interesting fossils, but they do not provide enough evidence to get the dino-bird hypothesis off the ground. In fact, the heavy tails and large bodies of theropod dinosaur bodies are the exact opposite of the type of organism we would expect to evolve into a flier92.
"These guys are mixed up; the description they refer to sounds like Sinosauropteryx (a compsognathid). No one has questioned the feathers on Protarchaeopteryx; the only question was whether or not it was a bird."
According to the report in Nature38, Protoarchaeopteryx, was a dinosaur with "down-like" feathers on its body, however one paleontologist called this down "dino-fuzz, [which] really could have nothing to do with the origin of feathers."91
Regardless, down feathers have no function for flight on birds. The only "vaned, barbed, symmetrical feathers"38 on Protoarchaeopteryx appear on its tail--no sign of feathers for flight. Caudipteryx has a few vaned and barbed remiges on one finger of the hand, but its arm is far shorter than that of a bird39--too short for flight. Because these organisms have some body structures which are well adapted to flight, but clearly didn't fly, these (and other) fossils have led evolutionists to believe that some of the primary complex structures specified for flying -- feathers, wings, and ossified bones -- originated for a purpose other that flight40.
"Neither of these guys have 'wings'; they are just arms. Those of Caudipteryx, in fact, are really short compared to other oviraptorosaurs!
"'ossified bones' = bones made of bones. Since you and I have ossified bones, as do cats, dogs, lizards, frogs, and trout, this is hardly a flight characteristic!!"
There are no elegant explanations here, but rather wishful thinking trying to force-fit the data to an evolutionary model failing to explain the origin of flight.
"Actually, what there is is the description of the material at hand. Fossils first, guys; hypotheses second."
"Nope. These have been shown to be structurally disctinct from the true feathers of birds and other dinosaurs; see recent papers by Reisz and Sues."
Reptiles of a feather, don't flock together
In fact, feathers have been also found a non-dinosaur lizard-like reptile90,
however proponents of the dino-bird hypothesis think this feathered fossil is completely unrelated to the origin of birds and the alleged feathered dinosaurs. Why, then, should the alleged appearance of feathers on these post-avian Dromaeosaurs become the definitive proof of the dinosaurian ancestry of birds? Perhaps these "feathered dinos" are just what their squamate
"Longisquama is NO squamate!!! (Squamate = lizards & snakes). It isn't a true archosaur, either, as it has traditionally been considered.
"Furthermore the primary evidence of the dinosaurian origin of birds is skeletal, not integumentary. See my website, among many other references."
"Identical."
counterparts are: reptile chimeras unrelated to birds.
To throw some other bones into the dino-bird hypothesis, although both theropod dinosaurs and birds both walk on 2 legs and have some skeletal similarities, differences such as digit configuration,
"identical (and in fact the same thing: pubis bone being part of the pelvis!) in basal dromaeosaurids, basal troodontids, and basal birds."
pubis bone, pelvis shape,
"Teeth of primitive birds, primitive troodontids, and primitive dromaeosaurids all very similar."
teeth,
"Fair to say that it is distinct from all LIVING reptiles."
and internal organ setup (the avian respiratory system is unique and far different than of reptiles)
And rejected...
have all been raised
as differences which challenge the dino-bird hypothesis43, 92. Wells also notes that cladistical methods which have established this alleged relationship ignore the fossil record,
"Bullcrap. How would a cladistic analysis of forms only known from fossils ignore the fossil record?
"(I suspect he meant 'stratigraphic record', but that is another issue)"
"Guilty. :-) "
assume an evolutionary history,
and ignore major problems with the implications that dinosaurs evolved to fly from the ground.
"????? Phylogenetic analyses are hypotheses of relationships; they are the framework by which other studies (like flight origins or so forth) might be interpreted, but the analyses themselves are JUST the interrelationships."
The full text of the Holtz email is here.
Enough of my creationist rhetoric--does Wells get it right?
With regards to moths, Tamzek quotes Majerus who, in response to Wells, admits that the melanism hypothesis is "discredited", which is indeed what is taught in textbooks, validating Wells' general critique. In this 2000 response to Wells, quoted by Tamzek, Majerus says that differential bird predation is still responsible for the changing coloration of the moths. Yet, this same Majerus wrote in 1987, "it seems certain that most B. betularia rest where they are hidden.... [and] that exposed areas of tree trunks are not an important resting site for any form of B. betularia"87, which would preclude the possibility that bird predation is an important factor in their selection. Majerus, an expert, seems to be either contradicting himself here, or retracting his claims.
In the quoted correspondence Majerus does not say that the idea that birds preying on differentially camouflaged moths results in industrial melanism was "discredited", he explicitly says there is substantial evidence for it. He did say that the account presented in textbooks is oversimplified, which does not validate Wells' critique.
If the author had actually read reference 87 (Howlett R.J., Majerus M., Biological Journal of the Linnean Society 30: 31-44 (1987)) he would have noted that Majerus and Howlett did not dispute that bird predation was an important factor in selection in that paper. Their concern was in getting accurate quantitative values for selection via bird predation. The author should have also noted that Howlett and Majerus reported the results of an experiment comparing selection pressure in exposed and unexposed locations. There was no significant difference in the ratio of dark to light moths taken by birds between exposed and unexposed locations, thus bird predation is an important factor in both locations. Majerus's stance in 1987 was the same as that in 2000.
-- Ian Musgrave
Regardless of what moths really do, Wells was not unjustified in trusting Majerus, an expert (and others88), when Majerus implied that birds are not a factor in moth selection87. Majerus and others may disagree with Wells, however Wells himself is not completely ignorant on the subject of moths, as he had an opinion article published on the subject entitled, "Second Thoughts about Peppered Moths"88 in the peer-reviewed journal, The Scientist.
The peer-review assertion is a false statement. The Scientist is described as "The News Journal for the Life Sciences" (http://www.the-scientist.com/). In a brief search of the website, I can't find anything mentioning peer-review or the lack thereof, nor any submission guidelines. This is not a good sign. In any case, Wells' piece was in the "Opinion" section, as you can see right here.
I have also emailed The Scientist for clarification regarding peer-review. The publisher, Alexander M. Grimwade Ph.D., responded:
1. THE SCIENTIST is NOT a peer reviewed journal.
2. The article written by Wells was published in our Opinion section, where we allow scientists to "sound off" with minimal editing or review of content. To quote our masthead:
"THE SCIENTIST serves its readers in many ways, but one of its most valued aspects is its committment to open discussion of controversial topics. While readers often praise us for this committment, they should also recognize that all articles published in THE SCIENTIST reflect the views of their authors and not the official views of the publication, its editorial staff, or its ownership."
That The Scientist may not have fully understood the moth situation when they accepted Wells' piece is indicated by their cartoon. Who would go moth-hunting in a lab coat and with a magnifying glass?
In addition, the original version of Luskin's response said that Wells' piece in The Scientist "contained over 45 references" -- actually it had 17. The expanded version of Wells' moth essay is available at ARN, which is probably what Luskin was referring to, although a careful count of the reference list reveals 44 references. In any case, the number of references is unimportant if one is using them unfairly, as I think the expanded version of peppered moth section of my article shows.
If, however, you are a real glutton for well-referenced detail regarding peppered moths and Wells, then the person to consult is Ian Musgrave. Some of his comments are online here.
-- Nic Tamzek
Majerus never implied bird predation was not an important factor in selection. In fact he explicitly states that it is, and emphasizes that his concern is for getting an accurate quantitative estimate of the selection pressure due to bird predation. That Wells was able to get published in "The Scientist" says nothing about his knowledge of moths. "The Scientist" is not a peer-reviewed publication, but a publication dedicated to general interest items and news. Any more than a superficial examination of the Howlett and Majerus paper would have shown that Majerus was not disputing that bird predation was an important (though not necessarily exclusive) factor in industrial melanism. Not that this is surprising, not even Kettlewell thought that predation was the only factor involved (Kettlewell, HBD. (1961) Ann. Rev. Ent., 6: 245-262.).
-- Ian Musgrave
Not to pile on here, but this point cannot be emphasized enough. Majerus repeatedly and clearly has stated his view that the textbook stories are oversimplified, but that the basic point, which is that bird predation of moths is the major selective force causing the change in moth color, remains a good conclusion well-supported by the data.
Here are Majerus' conclusions on the peppered moth case, at the end of the two chapters (nearly sixty pages) of his book Industrial Melanism that he spends precisely on this topic:
The case of the peppered moth is undoubtedly more complex and and fascinating than most biology textbooks have space to related. Even here, I have had to be selective about which data sets to include, given the huge wealth of material on this case. I have tended to concentrate on work carried out in the industrial northwest of England, because this is the most extensive work, and East Anglia where I have worked myself. My aim in this and the preceding chapter has been to provide sufficient information for the reader to assess for themselves which parts of the story are complete, and where further work is needed (see Chapter 10). My view of the rise and fall of the melanic peppered moth is that differential bird predation in more or less polluted regions, together with migration, are primarily responsible, almost to the exclusion of other factors. (Majerus, 1998, Industrial Melanism, p. 155, emphasis added)
-- Nic Tamzek
Anthropologist Jeffrey K. McKee, author of The Riddled Chain : Chance, Coincidence, and Chaos in Human Evolution, provided a reply to this via email:
Paleontologist Stephen Jay Gould would most likely concur with Wells as he once wrote, "[m]ost hominid fossils, even though they serve as a basis for endless speculation and elaborate storytelling, are fragments of jaws and scraps of skulls."48 Roger Lewin would also be in agreement when he wrote, "the major problem has been the pitifully small number of hominid fossils on which prehistorians could exercise their interpretive talents"75. Wells is not at all dishonest to claim that the paucity of fossils useful for investigating the alleged hominid ancestry of humans causes subjectivity among researchers. Wells' statements are in line with what Gould and Lewin have said, and are not negated by a reduced level of the relevance found in Well's quotation of Henry Gee.
If one is trying to establish evidence that humans evolved, then the truth of the matter is that paleoanthropologists have an embarrassment of riches. There are thousands of fossils by which to document the major trends over the past 4 million years, starting with the earliest bipedal creatures (Australopithecus) who reatained upper limbs for climbing, and only had a modicum of brain expansion as compared to living apes. Early Homo (i.e. Homo habilis) showed further increases in relative brain size, and whereas the limbs changed subtly, their limb proportions remained similar to that of apes. Homo erectus/ergaster took the trends further, with hominids reaching a modern body size and general bodily proportions, but not modern facial shape or relative brain size. Archaic Homo sapiens then show a mixture of features appearing at different times and places, with a fully modern suite of features appearing about 100,000 years ago.
If one is trying to decide "who begat whom" and establish an accurate phylogeny, or to deduce the proximate evolutionary causes for the origin and spread of particular features, then more fossils would help resolve many of the specific questions ... that is what Gee, Lewin, Holden, and Gould were referring to. Scientists want to discern the "who, when, where, and how "of evolution, not just the "if."
McKee:
Missing: Link
Of the precious little that is found, the bottom line there are basically 2 types of hominid fossils: those of the genus Homo and those of the genus Australopithecus. Everything of the genus Homo (Homo erectus, habilus, neanderthalis, sapiens, etc.) have skulls and body shapes which are very similar to modern humans--often to the point that they could be within the possible range of modern human genetic variation.
As noted above, Homo habilis had limb proportions similar to that of apes, but the morphology of a biped. Whereas many of the individual features of the skulls of subsequent members of the genus Homo may be within the modern human range, the entire package is markedly different. For example, much is made of the observation that Homo erectus cranial capacity is within the modern human range. Fine, but the brain size for body size ratio is outside the modern human range.
McKee:
Everything else from the genus Australopithecus (from which the famous fossil "Lucy" is derived), looks much more like a chimpanzee (Australopithecus means "southern ape") There are "robust" forms of Australopithecus which are not very chimp-like, but these also look absolutely nothing like Homo, and evolutionists do not believe they are on the human line.
Indeed, Australopithecus DOES look chimp-like in many respects. It also has a relative brain size, teeth, bones, and inferred posture and gait, derived more toward humans than chimps ... exactly as one would predict of a member of the hominid lineage that evolved shortly after the divergence from a common ancestor of the lines leading to humans and modern chimps.
McKee:
So what fossils are there to link the chimp-like members of Australopithecus with the fossils of the genus Homo, which basically represents modern humans? The answer is nothing. Between the two are great skeletal differences with regards to locomotion49, 51 and a large gap in brain size50, with nothing in between. The "missing link," it would seem, is still missing.
The intermediate nature of the Homo habilis post-cranial skeleton, as mentioned above, defies the above statement about locomotion. But where is the "large gap" in brain size? For example, Australopithecus africanus has an average cranial capacity of 440 cc (cubic centimeters), wheareas the average for Homo habilis averages 610 cc -- and their ranges overlap. No big jump there. More useful, however, is a measure of relative brain size: the Encephalization Quotient, or EQ. Basically, the EQ tells us how much more brain an animal has than a typical mammal of its body size. So a chimp, for example, has an EQ of 2.0 -- twice the brain expected for a mammal of a chimps size. Now let's look at average EQs for the major hominid fossil taxa and look for major gaps: Australopithecus afarensis: EQ=2.2,;A. africanus: EQ=2.5 (considerably more than a modern chimp, by the way); Homo habilis/rudolfensis: EQ=3.1; early Homo erectus: EQ=3.3; late Homo erectus: EQ=4.0; modern Homo sapiens: EQ=5.8.
Is the "missing link" still missing? Yes, as every time paleoanthropologists find one more fossilized link of the chain, creationists find two more missing links (i.e. between the link and its antecendent, and between the link and its descendent). At the current rate of discovery and publication of fossil hominids, the number of missing links should increase by an order of magnitude by 2050.
After providing quotes from a number of authorities, including Gee, noting the lack of hominid fossils and the subjective nature of the many phylogenetic trees of human evolution Wells is justified in asking "[g]iven the highly subjective nature of paleoanthropologoy--as acknowledged by its own practitioners--what can the field reliably tell us about human origins?" (Icons pg. 223). The answer, it would seem, is definitely not that we have unquestionably evolved from apes.
[Note: the following response to Luskin's "Missing: Link" section was written before McKee's response was provided.]
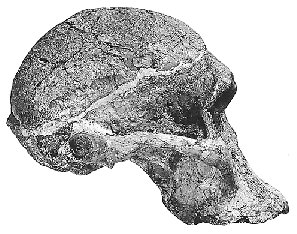
Luskin claims that fossil hominids can be broken up into two groups: the chimpanzee-like Australopithecus and Homo in which he includes Homo habilis saying they have "skulls and body shapes which are very similar to modern humans." This is breathtakingly wrong. While it is true that the australopithecines had many ape-like body characteristics, how many chimpanzees walk bipedially (on two legs) as the australopithecines did? And while the postcranial (i.e. not part of the skull) evidence for Homo habilis is meager, it is clear from the fossil evidence (in particular OH 62) that it had body shape similar to the australopithecines. Today what has been historically classified homo habilis, the habilines, is split into two species, H. habilis and H. rudolfensis, by many researchers. H. habilis includes a skull that no special creationist considers a human while H. rudolfensis has a skull that some creationists think is an ape and some think is a human and yet the two are very similiar. Recently Bernard Wood, an influential researcher, and Mark Collard published "The Human Genus" in Science (vol. 284, pp. 65-71, 2 April 1999) which they proposed that the both these two habiline species be reclassified as Australopithecus and not Homo. Various other researchers have previously proposed that some of the habiline fossils should be considered australopithecines. And Luskin wants us to believe that there is a clear line between Homo and Australopithecus? But I will let readers judge for themselves. The following table has some images of some skulls from Homo ergaster that had a rather modern body form, the two species of habilines, and an australopithecine species. The links below the images link to either a larger photo or more information about the particular fossil. The estimated cranial capacities (brain sizes) of each species are taken from the Wood and Collard paper.
| Species | Cranial Capacities |
FH = Fossil Hominids FAQ SI = Smithsonian Institution Human Origins Program |
||
|---|---|---|---|---|
| Homo Ergaster | 854 cm3 |  FH SI |
 FH SI |
|
| habilines | Homo (or Australopithecus) rudolfensis | 752 cm3 | 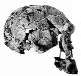 FH SI |
|
| Homo (or Australopithecus) habilis | 552 cm3 | 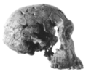 FH SI |
 FH SI |
|
| Australopithecus africanus | 457 cm3 |  FH SI |
 SI |
|
There are many other fossils as well. The reader is invited to go to this link and try to draw the line between "ape" and "human." It is not a simple matter matter as one would expect if ape and humans were separate creations as demonstrated Jim Foley's table of creationists' opinions of what the fossil hominids are.
-- Mike Hopkins
Wells is not a fraud
Apart from his doubts of Darwinism, a review of Tamzek's charges against Wells reveals that there is not one established instance where Wells is wrong, and few where he is even clearly at odds with mainstream scientific thought. The only bona-fida error in Icons could be found in Wells' figure 5-3 on the Cambrian explosion, which Tamzek doesn't verify, and this alleged error itself would not affect the strength of Wells' arguments about the Cambrian explosion. How is it that Wells can be seemingly be in agreement with mainstream science in almost all of his arguments (even Eugenie Scott says that Wells is "usually technically correct"89), which ought to have a profoundly negative effect upon the evidence for evolutionary theory, yet Wells differs from mainstream science in his conclusion that evolution might be false? The answer most likely lies in Wells' interpretation of the data he collects in analyzing the level of errors in textbooks.
The full text of Eugenie Scott's review can be found
at:
http://www.scienceormyth.org/icons%20of%20evolution.html
Luskin quotes her out-of-context as was pointed out here. This is a bit more of what she wrote:
...This is because Wells presents a systematically misleading view of evolution. Individual sentences in Icons are usually technically correct, but they are artfully strung together to take the reader off the path of real evolutionary biology and into a thicket of misunderstanding.... [Emphasis added.]
-- Mike Hopkins
Step into the light
In conclusion, Tamzek quotes Theodosius Dobzhansky, a theistic evolutionist, to show that one can believe in God and in evolution. In the quote Tamzek provides, Dobzhansky seems to be arguing only against a young-earth perspective, which is not at all what Wells advocates. In fact, Wells might even agree with Dobzhansky that religious texts are not "textbooks of astronomy, geology, biology, and anthropology". Where Wells would disagree, is that one's conclusion therefore ought to be that, empirically speaking, God had no direct effect upon the creation process, and that God created everything through undirected evolution. Biblical Scripture may say little about the details of the actual processes God used to create, however it is clear that God intended things to be as they are57. Dobzhansky would be hard pressed to fit the randomness of evolution into God's purposefulness. For the theist willing to dare to give up academic credibility by acknowledging counter-evidences to biology's sacred cow105, these problems can be solved. Had Dobzhansky lived to see the birth of the intelligent design movement, perhaps he too as a theist would have embraced its more open-ended approach.
At some fundamental level, all creationists believe generally in intelligent design. That is, however He chose to do it, all creationists believe that God created the world and its species, including man, purposefully. In his essay, "Nothing in Biology Makes Sense Except in the Light of Evolution," Dobzhansky explicitly called himself a creationist and an evolutionist, which by substitution means that he considered himself an advocate of evolutionary intelligent design or theistic evolution. (Henceforth, all references to "intelligent design" will be intended to refer to "anti-evolutionary intelligent design" and not theistic evolution. Nevertheless, it should be emphasized that, in general, there is no necessity for an intelligence to use strictly non-Darwinian processes to create the diverse lifeforms in this world. Theistic evolution is a legitimate position of intelligent design and, notably, it is only Wells or his ilk's version of anti-evolutionary intelligent design that makes an enemy of evolution or the methods of science.) Hence, trying to support the idea that Dobzhansky would advocate any form of a nti-evolutionary creationism, especially the modern articulations of intelligent design, would be quite a challenge.
In his response to Tamzek, Luskin attempts to pull off precisely this Herculean feat. Luskin tries to argue that Dobzhansky was ignorant of both the contributions of random chance to evolution and of modern intelligent design methods for explaining observations. If true, Luskin reasons that Dobzhansky might have supported modern intelligent design reasoning had he simply been given the chance. Unfortunately, these statements are not true and Luskin's claim of possible support from Dobzhansky is unlikely.
First, the intelligent design movement is not new. Its modern roots go back at least as far as Michael Polanyi, who was a contemporary of Dobzhansky. Even omitting Polanyi's modern contributions to the intelligent design literature, Reverend William Paley's "Argument from Design" first described the basic intelligent design argument, and Paley lived well before even Darwin.
At some basic level, intelligent design arguments claim that a certain phenomenon cannot be explained by science and therefore necessitate the existence of God. [2] Behe and Dembski's contributions to the intelligent design literature, modern sounding and esoteric as they may be, still rely upon Paley's same, flawed logic; only the phenomenon in question has changed through time. Therefore, the arguments from intelligent design have not meaningfully changed since Paley first articulated them. Since Dobzhansky did not support Paley's reasoning, he would not likely have supported Behe or Dembski's reasoning either.
There is also a high likelihood that modern intelligent design arguments are designed to sneak through the establishment clause of our Constitution. Indeed, modern intelligent design proponents have only removed the positive claims about the designer that Paley made: where Paley explicitly argued for God, modern intelligent design advocates argue for a more vague, officially unnamed "designer." Such argumentative sleight of hand does not really alter the logic of Paley's argument. Rather, it simply makes unclear the identity of the designer, the purposes of the designer's design, and other positive claims about design. Again, the argument of modern intelligent design advocates is not substantially different from the argument of premodern intelligent design advocates.
For Luskin to claim that Dobzhansky never had an opportunity to consider intelligent design methods or arguments is to deny either the long history of intelligent design creationism or the congruence between modern and premodern intelligent design arguments. Instead, it is likely that Dobzhansky presciently included them even as he made the case against anti-evolutionary creationisms in his essay that predated both Behe and Dembski.
Second, evolution involves, at least in part, a contribution from random chance -- it always has, ever since Darwin wrote his first edition of Origin. Thus, as a contributing scientist to the field of biology and the study of evolution, it is unlikely that Dobzhansky would have failed to consider randomness in the evolution that he thought was perfectly compatible with modern theology. Consider the following quotation from his essay.
The organic diversity becomes, however, reasonable and understandable if the Creator has created the living world not by caprice but by evolution propelled by natural selection. It is wrong to hold creation and evolution as mutually exclusive alternatives. I am a creationist and an evolutionist. Evolution is God's, or Nature's method of creation. Creation is not an event that happened in 4004 BC; it is a process that began some 10 billion years ago and is still under way.
An entire section of this essay is devoted to the discussion of organic diversity. There is little doubt that Dobzhansky credited randomness for that diversity, at least in part. However, the words "random" or "chance" do not appear in Dobzhansky's essay. Nevertheless, he is quite clear in other writings that evolution involves contributions from randomness and chance. Consider this quotation from Futuyma's textbook:
As Dobzhansky (1970) said, "It may seem a deplorable imperfection of nature that mutability is not restricted to changes that enhance the adaptedness of their carriers. However, only a vitalist Pangloss could imagine that the genes know how and when it is good for them to mutate." {Futuyma, Evolutionary Biology 2nd ed. 1986, p. 76}
Or consider this quotation from the American Scientist:
The process of mutation is the only known source of the new materials of genetic variability, and hence of evolution. {T. Dobzhansky, American Scientist, v. 45, 1957, p.385}
In the light of these statements, it would seem that Dobzhansky did indeed recognize randomness in his understanding of evolution. "Nothing in Biology Makes Sense Except in the Light of Evolution" was written in 1973 and the above quotations were written in 1970 and 1957. By any reasonable assessment, Dobzhansky included random influences in the evolution that he believed was compatible with theism.
Luskin suggests that Dobzhansky was ignorant of randomness in evolution, though Dobzhansky knew quite well of the randomness involved in evolution. Luskin argues that Dobzhansky was ignorant of modern intelligent design arguments, though Dobzhansky knew quite well the flaws in Paley's prototypic version of the same. Luskin's suggestion that Dobzhansky might have agreed with intelligent design creationism is therefore unsupported and, in fact, highly unlikely.
In contrast to Tamzek, Wells uses Dobzhansky's quotes for purely scientific purposes--to show that Dobzhansky's famous quote--"[n]othing in biology makes sense except in the light of evolution"104--represents a departure from the scientific method, as it presumes that everything in biology must fall under the evolutionary paradigm:
"[S]cience at its best pursues the truth. Dobzhansky was dead wrong, and so are those who continue to chant his antiscientific mantra. To a true scientist, nothing in biology makes sense except in light of the evidence." (Icons, pg. 248)
Dobzhansky wrote an essay entitled "Nothing in Biology Makes Sense Except in the Light of Evolution." Notably, Dobzhansky wrote an essay with this title; he did not write the title only. In the essay, Dobzhansky makes it clear why he titled his essay as he did: evolution explains observations better than any other explanation offered and biology without evolution would be a pile of observations without a unifying theme.
Consider the following excerpts from the essay in question:
The environment does not impose evolutionary changes on its inhabitants, as postulated by the now abandoned neo-Lamarckian theories.
All this is understandable in the light of evolution theory; but what a senseless operation it would have been, on God's part, to fabricate a multitude of species ex nihilo and then let most of them die out!
They fancy that all existing species were generated by supernatural fiat a few thousand years ago, pretty much as we find them today. But what is the sense of having as many as 2 or 3 million species living on earth?
Was the Creator in a jocular mood when he made Psilopa petrolei for California oil fields and species of Drosophila to live exclusively on some body-parts of certain land crabs on only certain islands in the Caribbean?
But what if there was no evolution and every one of the millions of species were created by separate fiat? However offensive the notion may be to religious feeling and to reason, the antievolutionists must again accuse the Creator of cheating. They must insist that He deliberately arranged things exactly as if his method of creation was evolution, intentionally to mislead sincere seekers of truth.
Notice how Dobzhansky is specifically demonstrating how he has considered other explanations, both non-natural and natural, but found them all lacking. Having considered those alternatives, which are likely representative of all forms of anti-evolutionary creationism or non-Neodarwinian evolution (Lamarckian evolution, etc.), Dobzhansky entitled his essay as he did. Had Dobzhansky not considered any alternatives, Luskin's objection would be accurate: there is a chance that another hypothesis of biodiversification would fit the data; therefore, considering evolution as the only possible explanation is either outright invalid or dogmatic.
Unfortunately for Luskin, the essay makes it quite clear that the title does not fully convey or develop Dobzhansky's ideas. It may be possible to explain data from a non-evolutionary perspective, but as Dobzhansky points out often in the essay, those perspectives are absurd and not nearly as sound or as useful as evolution. Thus, Dobzhansky's title remains valid and Luskin's stated point is easily dismissed.
There is another way that Luskin's objection might be valid, though he does not state it himself. What if evolution were easily removed from the field of biology en bloc? If scientists were somehow able to understand biology without evolution, then the suggestion implied by a literal reading of Dobzhansky's title -- that nothing in biology can possibly make sense without evolution -- would be inaccurate and Luskin's objection might be sustained.
Most likely, it was to this very objection that Dobzhansky wrote in his essay:
Seen in the light of evolution, biology is, perhaps, intellectually the most satisfying and inspiring science. Without that light it becomes a pile of sundry facts some of them interesting or curious but making no meaningful picture as a whole.
Dobzhansky is saying that the extrication of evolution from our modern understanding of biology would either be a difficult process or would render biology incomprehensible altogether. By any other language, he is saying that nothing in biology makes sense except in the organizing, explanatory "light" of evolution, a justifiable interpretation that also undermines Luskin's objection.
Creationists often try to undermine the logic of Dobzhansky's essay by claiming that there are indeed observations in biology that do not require an evolutionary perspective to comprehend. Wells seems to follow suit by also arguing against the title instead of the essay, though he prefers to attack by dismissing evolution as a mere paradigm. These arguments both presume that the reader has read only the title and not the essay that bears it. If one reads Dobzhansky's essay, the title choice is justified, even if it is a clunky summary of the essay itself. If Luskin is arguing that the title of Dobzhansky's essay does not develop Dobzhansky's essay as completely as the essay itself does, his argument is unremarkable and easily dismissed by reminding readers that an understanding of issues is seldom accomplished by judging proverbial books by their covers.
In truth, Luskin and Wells' points are both easily addressed by reading the essay in question [http://www.2think.org/dobzhansky.shtml]. The title of that essay is completely justified by the essay's development: it is neither a deviation from the scientific method, nor is it flawed in its reasoning.
With regards to the methods of science, how ironic that Wells, as an intelligent design creationist, would accuse Dobzhansky of having deviated from the scientific method. The answers to biological questions that intelligent design advocates advance, because those answers invoke unobservable and immeasurable "supernatural" bringers of natural change, are answers that are not testable by science. It is intelligent design creationists, and not Dobzhansky, who deviate from the scientific method -- and they deviate from it broadly. Hence, Wells cannot seriously accuse anyone of deviating from the methods of science without engaging in glaring hypocrisy.
Wells' point is that the evidence--not a paradigm--should be controlling our understanding of the data. After a look at weaknesses in the most common lines of evidence used to support evolutionary theory, at least 2 things are clear: evidence for evolutionary theory is surely lacking in some important places, and Icons of Evolution will be a royal pain in the fanny for evolutionists for years to come.
The current scientific understanding of the observable and measurable "natural" world is the one that explains observations and accurately predicts new data the best. New data and new theories are considered in the light of these current understandings. It is the process of science to continually reevaluate the status of considered explanations, keeping the theories that best explain and predict the data and de-emphasizing or outright rejecting the rest. In defining evolution as the current scientific understanding of biodiversification, one should infer that it therefore best explains and predicts findings from the testable world.
Wells recognizes that evolution has a broad theoretical framework to which new data are applied. However, he decries it as merely a "paradigm," as though evolution were a theory that is in principle not capable of being refuted by natural observations. In making this argument, he ignores the fact that from the day Darwin conceived it, evolution was capable of being refuted. Thus far, it hasn't been.
It is not a biased treatment for evolution or against intelligent design that keeps evolution the current scientific understanding for biodiversity and other observations. Far more than simply a "paradigm" to guide thought in biology or not, evolution is instead a workhorse of a scientific theory. It has guided countless experiments, it has explained so many observations and measurements in diverse fields, and has been verified and buttressed so completely that it is as close to a fact as science will likely ever get. It is truly "The integrating theory of biology."
Thus, Wells' suggestion that evolution is simply a favored "paradigm" or has no useful theoretical backbone is quite wrong. Evolution is instead the current scientific understanding because evidence has robustly supported it to be.
Regarding evidence, Wells has often styled himself as a biologist who simply disagrees with evolution. If Wells really acted like a scientist or actually utilized the training he learned in pursuit of his advanced biology degree, he would be working to develop the evidence needed to support his case against evolution. Instead, Wells argues against the definition of science, calling the restriction of science to testable, "natural" hypotheses biased. Wells probably recognizes the robust nature of evolution and, to the end of furthering his admitted crusade against Darwinism for which he sought advanced credentialing as a biologist in the first place, considers it easier to undermine the methods of science rather than scientifically support his hypothesis.
Rather than succumb to Wells' stratagem, the reader is invited to examine the evidence supporting both evolution and intelligent design and make a decision as a scientist would. The journal Evolution has 18+ rigorously peer-reviewed articles per month. Across all journals, several original research papers on evolution are published every day, and countless more studies besides are made possible only because of the insights evolution provides or predictions evolution makes. Contrarily, it is indeed remarkable that intelligent design has no such papers. They aren't even trying to do real research because they can't; no data can ever contradict a postulated supernatural agent and appealing to the unobservable and immeasurable actions of supernatural agents to explain natural data is simply not a falsifiable hypothesis in science.
Despite a complete absence of scientifically meaningful evidence, Wells and the other intelligent design creationists request that the "evidence" for intelligent design be allowed alongside evolution in science classrooms, on the basis that the evidence and not a "paradigm" should control our understanding of the data. If one really based the inclusion or rejection of science classroom discussions upon the evidence, then intelligent design would quite simply not appear in a science classroom. Wells' argument therefore becomes as hypocritical as his previous one: he has no evidence to back up his claim other than an appeal to ignorance.
Luskin, calling upon Wells' research, then asserts that there are "flaws" in evolutionary theory. His pejorative description of current research as "flaws" is certainly worthy of rebuke, which is forthcoming. First, this response must begin by reminding readers of the substance of Tamzek's original critique of Wells. Any unbiased investigator will recognize that Wells cannot be trusted to adequately convey the state of evolution to readers. Simply because Wells thinks certain lines of evidence in support of evolution are "flawed" does not mean that open minded observers would consider them flawed. Luskin would do well to find other support for his conclusions than the statements in Wells' book.
It is unfortunate, but revealing, that Luskin pejoratively describes active areas of research in evolution as "flaws." In doing so, he intentionally connotes illegitimacy of evolution even while evolution is proceeding through a glorious age of success and utility. Indeed, Luskin's use of the pejorative reveals more about his personal bias against evolution than he reveals about evolution. Clearly, there is a need for more research in the field of evolution; no one doubts this. However, more research is needed in evolution for the same reasons that more research is needed in chemistry. That is, there will always be more to learn about chemistry and some of what we think we know about chemistry will turn out to be wrong; however, this does not support alchemy.
The fact is, science is an active discipline and there is and will always be a frontier beyond which we have yet no understanding and at which our understanding is incomplete. At and near this frontier, our models are subject to modification, perhaps rejection, as new information becomes available. However, the existence of uncertainty at this frontier does not negate our knowledge in the well-trodden foreground. [1] With apologies to Einstein, "If we knew everything about [evolution], it wouldn't be science."
It is true that there is still much research needed about evolution: The details about the mechanisms of evolution, the particular paths it took in particular lineages, the application of evolution to other problems or data not necessarily related to speciation, etc. all need to be investigated and solved. However, the intent of Luskin's statement -- that pivotally "important flaws" exist in the theory of evolution and these "flaws" undermine evolution as a whole -- is invalid for at least two reasons.
First, it is important to understand that at the "well trodden foreground" of evolution fundamentals, it is solidly articulated with adequate -- nay, overwhelming -- scientific support. The debate about evolution or the research being done on evolution is not over whether it occurred but over how it occurred. Put into different language, at the level of education in which evolutionary concepts are actively being investigated, one would likely be enrolled in a graduate school course: high school- and undergraduate-level evolution is as close to factual as science can likely get. Quite simply, the areas of active research in evolution -- Luskin's so-called "flaws" -- are not of a kind that would cause the unbiased to doubt whether evolution occurred; the "frontiers" of evolution lie far afield of the established fundamentals, about which there is no serious scientific objection. Hence, Luskin is quite wrong to claim there are "flaws" in evolution, if his intent in citing those "flaws" is to suggest that they undermine evolution.
Second, notice Luskin's unstated assumption: a failure of our current understanding of evolution can be taken as a success for intelligent design. This logic suggests that intelligent design is a default explanation: rule out evolution and intelligent design is all that remains. His assumption is seriously flawed for at least two reasons. First, a common failure of an argument from ignorance is that overcoming the ignorance in question easily undermines it. In general, given the history of science and its glorious history of overcoming ignorance of phenomena, the gaps in our current understanding of evolution will be filled eventually. Specifically, Luskin's attempt to rule out current understandings of evolution as "flawed" does little to rule out future understandings of evolution.
Second and more importantly, what is it about intelligent design that makes Luskin think it deserves default status? The understanding of evolution that we currently have -- which has active areas of investigation in advanced evolutionary concepts -- has far more of a theoretical, predictive, explanatory, useful framework to it than does intelligent design creationism, which frankly has none of these things. To consider that "evidence for evolutionary theory is surely lacking in some important places" and to conclude that intelligent design is therefore a better explanation for perfectly natural observations is to abandon perspective, reason, or the methods of science.
As for Wells' book being a "pain in the fanny to evolutionists," indeed he has taken on the role of a Socratic gadfly, arousing response from the slumbering scientific and religious communities. Previously, these communities did not regard creationism as a threat. Following the publication of Wells' book, they became more active in addressing creationism.
As a result, at any location in which Wells or intelligent design creationists now speak, scientists and theologians rebut their rhetoric with an interest and passion that they would not have otherwise had. This rebuttal of Luskin, for example, has been created with the assistance of many contributors, some of them professionals in the field who don't usually participate in creationist debates. In Kansas, and likely in other states as well, there is an interest in evolution that was not present before Icons was released. By any measure, Wells' book has done more to further evolution than it has done to damage it.
The reader is also again reminded of Tamzek's original rebuttal of Wells, in which the egregious misrepresentation of science by Wells is exposed. As Wells has failed to demonstrate academic integrity, the publication of his book has or will come at great personal cost: Wells will not be considered a reputable investigator or author by any group other than the intelligent design faithful.
Considering the blessings to evolution paradoxically provided by Icons and the costs to Wells' credibility, if the publication of Icons as a "pain in the fanny to evolutionists" is a victory for Wells, it is both a Pyrrhic victory for him and one that science and religion both are glad to concede.
Endnotes:
#1: Abell, GO (1983). "The Ages of the Earth and the Universe." Scientists Confront Creationism: New York: WW Norton & Company.
#2: Pennock, Robert T. (2000) Tower of Babel: The Evidence against the New Creationism. Cambridge: MIT Press. P. 163.
-- Burt Humburg
Note: This document was edited by several people including Nic Tamzek and Mike Hopkins. The original introduction was written by Nic though several others have made changes to it.

Home Page | Browse | Search | Feedback | Links
The FAQ | Must-Read Files | Index | Creationism | Evolution | Age of the Earth |
Flood Geology | Catastrophism |
Debates
{kind=link}