Alles über Archaeopteryx
von Chris Nedin[Referenzen bearbeitet: 10. Juni 2002]
![]()
Inhalt
- Einleitung
- Archaeopteryx-Fundstücke
- Archaeopteryx-Merkmale
- Archaeopteryx + moderne Vögel, nur Variation innerhalb einer Art?
- Ratiten
- Die Geschichte von zwei Becken
- Fliegen oder nicht fliegen
- Die Abstammung des Archaeopteryx
- Protoavis
- Fazit
- Danksagungen
- Literatur
Einführung
Archaeopteryx lithographica ("antikes Flügel aus dem Druckstein").
Nach dem Kalkstein benannt, in dem es entdeckt wurde. Der Stein ist ein glatter, feinkörniger Kalkstein, der in der Druckerei verwendet wurde. Aus dem und um den Solnhofener Bereich in Deutschland abgebaut. Gebildet am Boden einer hypersalinen Lagune im späten Jura, vor etwa 150 Millionen Jahren.
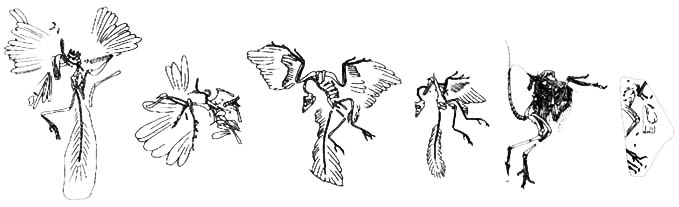
Archaeopteryx-Exemplare
Es wurden 8 Exemplare von Archaeopteryx gefunden (7 tatsächliche Exemplare und eine Feder). Diese Funde sind chronologisch (nach Beschreibung) unten dokumentiert.
- Die Feder
Entdeckt 1860 in der Nähe von Solnhofen und eine Offenbarung, als sie 1861 von H. v Meyer beschrieben wurde. Die Überraschung war nicht das Alter des Fossils, da mehrere Ornithopoden-Dinosaurier-Fußabdrücke, die fälschlicherweise Vögeln zugeschrieben wurden, bereits aus dem Trias bekannt waren, sondern die erhaltene Detailtreue.
- Das Londoner Exemplar

Entdeckt 1861 in der Nähe von Langenaltheim. Wahrscheinlich das bekannteste Exemplar (zusammen mit dem Berliner Exemplar). Seine Entdeckung wurde 1861 von H. v Meyer angekündigt, und das Exemplar wurde daraufhin vom British Museum of Natural History in London erworben (auf Anweisung von Richard Owen). Es kostete 700 britische Pfund – eine kleine Vermögen zu jener Zeit, aber für diesen Preis erhielt Owen auch etwas mehr als tausend weitere Fossilien aus Solnhofen. Das Exemplar wurde vom Amateur-Sammler und lokalen Arzt Carl Haberlein verkauft, der es als Zahlung für medizinische Behandlungen erhalten hatte. Owen beschrieb das Exemplar 1863. Er erkannte sofort, dass es sich um einen wichtigen Fund handelte, und stellte fest, dass es eine Übergangsform darstellt – jedoch nicht im Sinne von „Darwin". Owen war zwar ein überzeugter „Evolutionist", glaubte jedoch nicht an Darwins Modell der Evolution. Interessanterweise versäumte es Huxley, der ein überzeugter „Darwinist" war, die wahre Bedeutung des Fossils zu erkennen und beschränkte sich lediglich darauf, es als „vogelähnliches Reptil" zu bezeichnen. Erst durch detaillierte Vergleiche mit dem Dinosaurier Compsognathus wurde der wahre Wert des Archaeopteryx erkannt. - Das Berliner Exemplar

Entdeckt 1877 in der Nähe von Blumenberg. Dies war ein besseres Exemplar als das Londoner, hauptsächlich, weil es einen vollständigen Kopf besaß, obwohl dieser stark zerdrückt war, und vom Berliner Museum sofort erworben wurde. Es wurde von Carl Haberleins Sohn an sie verkauft (man könnte sagen, es blieb in der Familie!). Es wurde 1884 von W. Dames beschrieben.
- Das Maxburger Exemplar

Entdeckt 1958 in der Nähe von Langenaltheim (gleicher Fundort wie das Londoner Exemplar). Dieses Exemplar besteht nur aus dem Rumpf und ist das einzige, das sich noch in privater Hand befindet. 1992, nach dem Tod seines Entdeckers und Besitzers Eduard Opitsch, stellte sich heraus, dass das Exemplar verschwunden war, und es wird angenommen, dass es heimlich verkauft wurde (Abbott 1992). Sein Verbleib ist unbekannt. Das Exemplar wurde 1959 von Heller beschrieben.
- Das Haarlemmer oder Teyler-Exemplar

Dieses Exemplar wurde tatsächlich 1855 in der Nähe von Reidenburg entdeckt, also 5 Jahre vor der Feder! Es lag in einem Museum, nachdem es 1875 von H. v Meyer als Pterodactylus crassipes klassifiziert worden war. Merkwürdigerweise beschrieb Mayer es als über eine Flugmembran verfügend, die bei keinem anderen bekannten Pterodaktylus bekannt war; jetzt wissen wir, warum! Eine erneute Untersuchung des Fossils 1970 durch Ostrom enthüllte Federn und seine wahre Identität.
- Das Eichstätter Exemplar

Entdeckt 1951 in der Nähe von Workerszell, wurde es 1974 von P. Wellnhofer beschrieben. Dies ist das kleinste aller Exemplare, etwa zwei Drittel der Größe der anderen. Es unterscheidet sich auch in anderen Aspekten, wie der Zahnstruktur und den schlecht verknöcherten Schulterknochen. Es wurde vorgeschlagen, dass dies eine separate Gattung ist; jedoch können die Unterschiede auch auf die mögliche juvenile Entwicklungsstufe des Tieres und/oder eine andere Ernährungs-Nische zurückgeführt werden. Dieses Exemplar besitzt jedoch den am besten erhaltenen Kopf, von dem aus die Litanei der reptilen Schädelmerkmale des Archaeopteryx beschrieben wurde. Derzeit befindet es sich noch innerhalb der Gattung A. lithographica.
- Das Solnhofener Exemplar

Entdeckt in den 1960er Jahren in der Nähe von Eichstatt von einem türkischen Arbeiter. Zuerst als Compsognathus identifiziert von einem Amateur-Sammler, zeigten jedoch weitere Untersuchungen, dass die Arme für die Körpergröße zu lang waren, und die Präparation enthüllte Federreste. Beschrieben von P. Wellnhofer 1988.
- Das Solnhofen-Aktien-Verein-Exemplar
Ein neues Exemplar wurde von Wellnhofer (1993) beschrieben, jedoch ist die Beschreibung auf Deutsch, sodass die Informationen begrenzt sind. Das Exemplar wurde als neue Art Archaeopteryx bavarica klassifiziert und wurde als besitzend eines kleinen verknöcherten Sternums sowie Federabdrücke gemeldet.
Archaeopteryx-Merkmale
In pseudowissenschaftlichen Kreisen wurde viel über die Stellung von Archae im evolutionären Schema gemacht. Das übliche „Argument" lautet, dass Archae kein Übergangsfossil zwischen Vögeln und Dinosauriern sein kann, weil es ein Vogel ist. Diese vereinfachte Linie verkennt die Tatsache, dass Archae zwar tatsächlich als Vogel klassifiziert wurde, dies jedoch auf der Grundlage von 4 Hauptmerkmalen geschehen ist – wovon 2 nicht einzigartig für Vögel sind. Diese Klassifizierung ignoriert die Tatsache, dass Archae zahlreiche Merkmale aufweist, die einzigartig sind, einzigartig darin, dass sie nicht von Vögeln besessen werden. Die avianischen Affinitäten von Archae sind zulässig auf der Grundlage der folgenden 4 Hauptmerkmale:
Die vogelartigen Merkmale des Archaeopteryx
- 1) Federn.
-
Federn sind das diagnostische Merkmal moderner Vögel. Dies ist eines der Hauptkriterien zur Einordnung von Archae als Vogel, da kein anderes modernes Tier Federn besitzt. Die Besessenheit von Federn ist ein Merkmal von Vögeln, also streichen Sie einen für die Vögel. Allerdings konnte Ende 1996 eine Entdeckung in China diese Sichtweise ändern. Ein kleiner Theropoden-Dinosaurier Sinosauropteryx (Chen et al. 1998) wurde mit dem, was wie Federn entlang des Rückens erhalten zu sein scheinen, gefunden. Die Identifizierung der Strukturen ist jedoch zweifelhaft (z. B. Unwin 1998), und einige zweifeln daran, dass die Strukturen Federn sind.
STOP PRESS
GeFederte Dinosaurier gefunden
Zwei Dinosaurier-Arten wurden kürzlich in Nordost-China gefunden, die Federn besitzen (Qiang et al. 1998). Protoarchaeopteryx robusta und Caudipteryx zoui zeigen Regiges, Rectrices und plumulöse Feder Eindrücke. Ferner sind sie keine Vögel, da ihnen ein umgekehrter (rückwärts gerichteter) großer Zeh (siehe Nummer 2 unten) und ein quadrratojugal Squamosal-Kontakt fehlt, wobei ein Quadrojugal durch ein Ligament mit dem Quadrat verbunden ist und ein reduzierter oder fehlender Prozess des Ischiums. Diese und andere Merkmale gruppieren Protoarchaeopteryx und Caudipteryx mit maniraptoran Coelurosauriern statt Vögeln.
[Systematik Hinweis (von Padian 1998): Systematiker definieren die Namen von Organismen nach ihrer Abstammung, in diesem Fall bestehen Vögel (Aves) aus Archaeopteryx plus lebenden Vögeln und allen Nachkommen ihres jüngsten gemeinsamen Vorfahren. Vögel werden diagnostiziert durch einzigartige Merkmale, die nur sie besitzen und die von diesem gemeinsamen Vorfahren vererbt werden. Selbst wenn Federn von einer breiteren Gruppe geteilt werden als nur Vögel, werden Vögel immer noch als Archaeopteryx und spätere Verwandte definiert. Protoarchaeopteryx und Caudipteryx sind keine Vögel, obwohl sie Federn besitzen, weil die Suite morphologischer Merkmale, die sie besitzen, sie als dazugehörig markieren maniraptoran Coelurosaurier Dinosaurus.]
Es scheint, dass Federn nicht mehr als ein einzigartiges Merkmal von Vögeln verwendet werden können.
Für eine Diskussion der gefiederten Dinosaurier gehen Sie zu Jeff Poling's Dinosauria page.
- 2) Gegengängiger Hallux (großer Zeh).
-
Dies ist auch ein Merkmal von Vögeln und nicht von Dinosauriern. Obwohl gegengängige große Zehen in anderen Gruppen gefunden werden, sind sie, so weit mir bekannt ist, nicht in Dinosauriern gefunden. Ein umgekehrter großer Zeh wird jedoch bei einigen Dinosauriern gefunden, und der Zustand wird bei einigen Theropoden-Dinosauriern angenähert.
- 3) Furcula (Wischbein) gebildet aus zwei Schlüsselbeinen, die in der Mittellinie verschmolzen sind.
-
Nun beginnen wir auf wackligem Boden. Es wurde früher angenommen, dass die Besessenheit einer Furcula Vögel von Dinosauriern unterschied. Tatsächlich waren bis vor kurzem selbst Schlüsselbeine selten und weit entfernt, selbst bei Theropoden-Dinosauriern (der vorgeschlagenen engsten Gruppe zu den Vögeln, aus der die Vögel hervorgegangen sind - siehe Ostrom 1976). Allerdings wurde gefunden, dass Theropoden-Dinosaurier tatsächlich Schlüsselbeine besaßen (z. B. Bryant & Russell 1993) und sie wurden bei mehreren Arten gefunden, z. B., Segisaurus, Velociraptor, Euparkeria, Ornithosuchus, Saltoposuchus, Ticinosuchus. Auch Chure & Madson (1996) berichteten Furculae in einem nicht-maniraptoran, Allosaurid-Dinosaurier.
Es wurde gefunden, dass die Schlüsselbeine oft klein und schlecht ossifiziert sind. Das ist keine Überraschung, da sie für Ihren durchschnittlichen Theropoden-Dinosaurier kaum evolutionären Vorteil bieten. Allerdings zeigen auch Vögel diese Variation in der Ossifikation, besonders unter den Karnivoren und einigen Papageien, sind Schlüsselbeine reduziert oder sogar fehlend. Daher kann die scheinbare Abwesenheit von Schlüsselbeinen bei einigen Theropoden- Dinosauriern gut auf schlechte Ossifikation zurückzuführen sein, anstatt auf wahre Abwesenheit. Allerdings wurden Furculas bei einigen Theropoden-Dinosauriern gefunden, nämlich den Oviraptorosauria (Barsbold et al. 1990, Bryant & Russell 1993), zum Beispiel Oviraptor und Ingenia. Somit erscheinen Furculas nicht als diagnostisch für Vögel und bestimmte Mitglieder der vorgeschlagenen engsten Gruppe zu den Vögeln scheinen nun Furculas zu besitzen, so dass es ein neutrales Merkmal ist.
Eine häufig zitierte Kritik daran ist, dass die meisten der hier aufgeführten Theropoden-Dinosaurier nach Archae datieren. Allerdings wird keiner davon als Vorfahre beansprucht, und Eupakeria ist eine Trias-Form. Das Vorhandensein von Schlüsselbeinen zeigt, dass dieses Merkmal ein Merkmal von Theropoden-Dinosauriern ist und daher wahrscheinlich bei frühen Theropoden vorhanden war.
- 4) Pubis verlängert und nach hinten gerichtet.
-
Dies ist ein Merkmal von Vögeln, aber es ist auch ein Merkmal einiger Theropoden-Dinosaurier, so dass es nicht diagnostisch für Vögel ist - ein weiteres neutrales Merkmal. Allerdings teilen die Pubis-Schafte von Archaeopteryx und Dromaeosauriern (eine Gruppe von Theropoden-Dinosauriern, die eng mit Vögeln verbunden zu sein scheinen) eine plattenartige, leicht gewinkelte transversale Querschnitt, der bei keinem anderen Archosaurier gefunden wird.
Reptilienmerkmale des Archaeopteryx
- 5) Prämaxilla und Maxilla sind nicht hornbedeckt.
-
Das ist gehobene Sprache für „besitzt kein Schnabel." Die Prämaxilla besitzt keine keratinisierte Hülle, daher hat Archaeopteryx keinen Schnabel. Der Schnabel entsteht durch den Prozess der 'Keratinisierung', der die Schleimschicht der Epidermis (Romanoff 1960) einbezieht und dessen Bildung somit unabhängig von der Kieferknochenbildung ist.
- 6) Wirbel im Stammbereich sind frei.
-
Bei Vögeln sind die Wirbel des Rumpfes immer verschmolzen.
- 7) Knochen sind pneumatisch.
-
D.h. sie scheinen Luftsäcke zu besitzen, wie dies bei Vögeln und bei einigen Dinosauriern der Fall ist (z.B. Witmer 1990, Brooks 1993). Es sollte hervorgehoben werden, dass frühere Behauptungen, wonach die Knochen von Archae nicht pneumatisch seien (Lambrecht 1933; de Beer 1954), auf fehlende Belege beruhten, d.h. dass die Knochen keine pneumatischen Poren aufweisen (durch die die Luftsäcke in die Knochen eintreten) und die Knochen keine der Vollständigkeit und Wölbungen zeigen, die die pneumatischen Knochen moderner Vögel auszeichnen. Britt et al. (1998) fanden Belege für das Vorhandensein von pneumatischen Knochen in Archaeopteryx:
"Hier untersuchen wir zwei Exemplare von _Archaeopteryx_. Diese Exemplare zeigen Belege für Wirbelpneumatizität in den Hals- und den vorderen Brustwirbeln, wodurch die phylogenetische Kontinuität zwischen den pneumatischen Systemen nicht-aviarer Theropoden und lebender Vögel bestätigt wird" (Britt et al. 1998, S. 374)
8) Schambeine mit einer plattenförmigen und leicht geneigten queren Querschnittsform-
Ein Merkmal, das mit Dromaeosauriern geteilt wird, aber nicht mit anderen Dinosauriern oder Vögeln
- 9) Die Gehirnhälften verlängern sich, werden schlank und das Kleinhirn befindet sich hinter dem Mittelhirn und überlagert es von hinten nicht oder drückt nicht darauf.
-
Dies ist wieder ein reptilienartiges Merkmal. Bei Vögeln sind die Großhirnhemisphären kräftig, das Kleinhirn ist so stark vergrößert, dass es sich nach vorne über das Mittelhirn erstreckt und dieses nach unten zusammendrückt. Daher ist die Form des Gehirns nicht wie bei modernen Vögeln, sondern eher eine Zwischenstufe zwischen Dinosauriern und Vögeln (z. B. Alexander 1990).
- 10) Der Nacken verbindet sich mit dem Schäkel von hinten wie bei Dinosauriern, nicht von unten wie bei modernen Vögeln.
- Der Ansatzpunkt der Halsbefestigung (von unten) ist bei Vögeln charakteristisch; _Archaeopteryx_ besitzt dieses Merkmal nicht, sondern stimmt mit theropoden Dinosauriern überein:
"Beachten Sie, dass dieser coelurosaurähnliche Hals bei _Archaeopteryx_ von hinten am Schädel nach hinten verlief – wie es bei Coelurosauriern [theropoden Dinosauriern] der Fall ist – und nicht von unten, wie bei späteren Vögeln." (Ostrom 1976, S. 137).
Schädel und Gehirn von Archae sind im Grunde reptilisch und sind nicht „vollständig vogelartig" (im Gegensatz zu einer bestimmten creationistischen Behauptung).
- 11) Die Wirbelkörperzentren der Halswirbel weisen einfache konkave Gelenkflächen auf.
-
Dies entspricht dem Archosaur-Muster. Bei Vögeln sind die Wirbel unterschiedlich; sie besitzen eine sattelförmige Oberfläche:
"Das auffälligste Merkmal der Wirbel sind die einfachen scheibenförmigen Gelenkflächen ihrer Wirbelkörper, ohne jegliche Anzeichen der sattelförmigen Gelenkverbindungen, die bei anderen Vögeln zu finden sind" (de Beer 1954, S. 17).
- 12) Langer knöcherner Schwanz mit vielen freien Wirbeln bis zur Spitze (kein Pygostyl).
-
Vögel haben einen kurzen Schwanz, und die Schwanzwirbel sind verschmolzen, um den Pygostyl zu bilden.
- 13) Die Prämaxilla- und Maxillaknochen tragen Zähne.
-
Keine moderne Vogelart besitzt Zähne (z. B. Romanoff 1960; Orr 1966, S. 113). Vogelembryonen bilden Zahnanlagen aus, produzieren aber keine tatsächlichen Zähne. Einige Vögel bilden später Rippen im Schnabel, doch besteht kein Zusammenhang zwischen diesen und den embryonalen Zahnanlagen, da die Rippen auch in anderen Bereichen des Schnabels entstehen, in denen zuvor keine Zahnanlagen gebildet wurden. Einige Vögel bilden hakenartige Strukturen, die Papillen sind und scheinbar mit dem Prozess der Keratinisierung des Schnabels zusammenhängen (Romanoff 1960), und haben nichts mit Zähnen zu tun. Sie besitzen keine Blutgefäß- oder Nervenzusammenhänge und produzieren kein Dentin.
Die Ausbildung von Zahnanlagen im Vogelembryo hat eine einfache evolutionäre Erklärung, da sie darauf hindeutet, dass die Vorfahren moderner Vögel Zähne besessen haben und dieses Merkmal bei modernen Vögeln unterdrückt wurde. Das Vorhandensein von Zahnanlagen in den Embryonen von Organismen, die im Erwachsenenalter keine Zähne besitzen, stellt für Anti-Evolutionisten ein Problem dar, da sollte ein Merkmal exprimiert werden, das im Organismus nie genutzt wird? Einige fossile Vögel zeigen eine Reduktion der Anzahl der Knochen, die Zähne besitzen. Sowohl Hesperornis als auch Baptornis besitzen keine Zähne auf der Prämaxilla (Archaeopteryx und theropode Dinosaurier besitzen Zähne sowohl auf der Maxilla als auch auf der Prämaxilla). Nicht nur das, Hesperornis besitzt einen Schnabel, jedoch nur auf dem Oberkiefer (Gingerich 1975). Es besitzt also die Hälfte eines Schnabels und Zähne. Ein gutes Beispiel für eine morphologisch intermediäre Struktur zwischen zahnbesetzten Vögeln, die keinen Schnabel besitzen, und schnabelbesetzten, zahnlosen Vögeln.
- 14) Rippen schlank, ohne Gelenke oder uncinale Fortsätze und verbinden sich nicht mit dem Brustbein.
-
Vögel haben kräftige Rippen mit uncinaten Fortsätzen (Stützen zwischen ihnen) und setzen sich mit dem Brustbein zusammen.
- 15) Beckengürtel und Femur-Gelenk sind archosaurisch statt avian (außer dem nach hinten zeigenden Os pubis, wie oben erwähnt).
-
Hier zeigt Archae wirklich seine Übergangscharakteristik. Während das Becken als Ganzes im Wesentlichen frei ist und den Beckengürteln der Archosaurier ähnelt, zeigt das Os pubis nach hinten – ein Merkmal, das Vögeln und einigen anderen vogelähnlichen Theropoden-Dinosauriern gemeinsam ist.
Was interessant ist, ist, dass beim Vogelbecken:
"Das Os ischii liegt unter dem hinteren Teil des Os ilium und darunter liegt wieder das Os pubis, das nach hinten gerichtet ist (d. h. so: =). Embryologische Studien zeigen, dass die besondere Position dieser Knochen das Ergebnis einer sekundären Rotation ist und dass der pectineale Fortsatz vor dem Acetabulum nicht das wahre Os pubis ist, wie einige Forscher behauptet haben." (Bellairs & Jenkin 1960, S. 258).
Anders ausgedrückt sieht das embryonale Vogelbecken, wenn es zuerst gebildet wird, in Form und Winkel zwischen Os ilium und Os pubis (45 Grad) sehr ähnlich dem "A"-förmigen Becken von Archaeopteryx (d. h. so: <) (z. B. Romanoff 1960). Das vollständig ausgebildete Becken mit allen Knochen parallel liegend ist das Ergebnis einer sekundären Rotation des Os pubis von "<" zu "=". Dies unterstützt die Ansicht, dass Vögel einen Vorfahren mit einem saurischischen Becken hatten, wie es der Typus besaß, den Archaeopteryx und andere Theropoden-Dinosaurier besaßen. (siehe auch Die Geschichte von zwei Becken unten)
- 16) Das Kreuzbein (die zur Befestigung des Beckengürtels entwickelten Wirbel) nimmt 6 Wirbel ein.
-
Dies ist das gleiche wie bei Reptilien und insbesondere bei ornithopoden Dinosauriern. Das Vogelsakrum bedeckt zwischen 11-23 Wirbel! So ist zwar die Variation, die bei modernen Vögeln beobachtet wird, groß, sie ist jedoch bei weitem nicht so zahlreich wie bei Archaeopteryx
- 17) Metakarpalien (Hand) frei (außer 3. Metakarpal), Gelenk Handgelenk-Hand flexibel.
-
Das ist wie bei Reptilien. Bei Vögeln sind die Metakarpalien mit den distalen Carpalia im Carpo-Metacarpus verschmolzen, Handgelenk/Hand verschmolzen. Alle modernen Vögel haben einen Carpo-Metacarpus, alle fossilen Vögel haben einen Carpo-Metacarpus – mit Ausnahme eines (raten Sie mal!) :-). Allerdings zeigen die Carpalia mehrerer Coelurosaurier-Dinosauriergruppen einen Trend zur Verschmelzung, und bei der späten Kreide-Form Avimimus ist ein echter Carpo-Metacarpus gebildet.
Es wurde vorgeschlagen, dass der Strauß und/oder andere Ratiten ebenfalls unfreie Handgelenk/Handknochen besitzen. Das ist nicht korrekt:
"Der Strauß, Emus, Ruderhühner, Kasuare und Kiwis werden oft gemeinsam als Ratiten bezeichnet, obwohl sie möglicherweise nicht eng miteinander verwandt sind. Sie haben winzige Flügel und können nicht fliegen, aber die Knochen ihrer Hände sind auf dieselbe eigenartige Weise wie bei fliegenden Vögeln verschmolzen, was darauf hindeutet, dass sie von fliegenden Vögeln abstammen." (Alexander 1990, S. 435).
Einige Ähnlichkeiten zwischen der Hand des Straußes und einigen der abgeleiteteren Theropoden-Dinosaurier wurden einst genutzt, um zu behaupten, dass die Ratiten 'ursprünglich' seien und sich vor dem Aufkommen des Fluges bei Vögeln entwickelt hätten. Tucker (1938b) zeigte jedoch, dass solche Ähnlichkeiten völlig oberflächlich sind.
"Er hat auf die vogelartigen Merkmale der Hand des Dinosauriers Ornitholestes hingewiesen als Beweis dafür, dass eine vogelartige Hand unabhängig vom Flug entwickelt werden kann, doch der Autor hat in der oben erwähnten Mitteilung [Tucker 1938b] darauf hingewiesen, dass die Ähnlichkeit völlig oberflächlich ist und dass die eigenartige Biegung und terminale Verschmelzung der Metakarpalia 2 und 3, die sowohl die Carnaten- als auch die Ratiten-Hand kennzeichnen, im Dinosaurier in keiner Weise [sic?] reproduziert werden." (Tucker 1938a, S. 334).
"Zurückkehrend nun zu den Gründen, auf denen man die Ansicht begründet hat, dass die Ratiten ursprüngliche Vögel waren, deren Vorfahren nie geflogen sind: Erstens wurde die Ähnlichkeit zwischen der Hand des Straußes und der des Dinosauriers als ungültig abgetan. Tucker (1938b) hat gezeigt, dass die Ähnlichkeiten, die zwischen ihnen bestehen, nur oberflächlich und ohne Bedeutung sind." (de Beer 1956, S. 65).
- 18) Nasenöffnung weit vorne, vom Auge durch eine große Präorbitalfenestra (Öffnung) getrennt.
-
Das ist typisch für Reptilien, aber nicht für Vögel. Wo bei Vögeln eine Fenestra vorhanden ist, ist sie immer stark reduziert und beteiligt an der Prokinese (Bewegung des Schnabels).
- 19) Der Deltoidkamm des Humerus zeigt nach vorne, ebenso wie die Radial- und Ulnarkondylen.
-
Typisch für Reptilien, aber nicht bei Vögeln anzutreffen
- 20) Krallen an 3 nicht verschmolzenen Fingern.
-
Kein modernes erwachsenes Vogel hat drei Krallen, noch besitzen sie unfusionierte Finger. Der juvenile Hoatzin und Tukanen haben zwar zwei Krallen, verlieren diese jedoch, wenn sie wachsen, der Strauß scheint seine zwei Krallen bis ins Erwachsenenalter zu behalten, aufgrund der frühen Beendigung der Entwicklung (siehe Abschnitt zu Ratiten). Im Fall des Hoatzins wird angenommen, dass diese Krallen dem juvenilen Vogel das Klettern ermöglichen. Es wurde behauptet, dass, da diese Vögel Krallen haben, sogar im juvenilen Stadium, das Vorhandensein von Krallen nicht als reptilisches Merkmal verwendet werden kann. Das ist jedoch nicht der Fall. Tatsächlich zeigen fast alle Vögel Krallen, jedoch im embryonalen Stadium, und sie gehen verloren, bevor der Vogel das Ei verlässt. Im Fall der wenigen, die Krallen bis ins juvenile Stadium behalten, ist dies lediglich die Ausdehnung des Zustands in das postembryonale Stadium. Wie McGowan (1984, S. 123) sagt:
"Indem es ein primitives reptilisches Merkmal behält, das andere Vögel kurz vor dem Verlassen des Eies verlieren [der Hoatzin], zeigt es uns seine reptilische Abstammung. Fern von Beweisen im Gegenteil ist der Hoatzin zusätzlicher Beweis für die reptilische Abstammung der Vögel."
- 21) Das Wadenbein ist im Bein gleich lang wie das Schienbein.
-
Dies ist wieder ein typisches Merkmal von Reptilien. Bei Vögeln ist die Fibula verkürzt und reduziert.
- 22) Metatarsalia (Fußknochen) frei.
-
Bei Vögeln sind diese zu einem Tarsometatarsus verschmolzen. Bei modernen Vogelembryos sind die Fußknochen jedoch zunächst getrennt, wie beim Erwachsenen Archaeopteryx, und dies ist ein weiteres Merkmal, das eine reptilische Abstammung für Vögel unterstützt. Schließlich, warum sich die Mühe machen, getrennte Knochen im Embryo zu produzieren und sie dann zu verschmelzen? Warum nicht von Anfang an eine verschmolzene Masse produzieren? Kein erwachsener moderner Vogel hat getrennte Metatarsalia, aber sie sind im Embryo zunächst getrennt. Dies kann im Rahmen der Evolution erklärt werden – Vögel entwickelten sich aus einer Gruppe, die unfusionierte Metatarsalia hatte.
Ceratosaurier, Avimimus und Elmisauridae zeigen alle echte Tarsometatarsen. Archae zeigt lediglich den Anfang dieser Struktur.
- 23) Gastralien vorhanden.
-
Gastralia sind „ventrale Rippen", Elemente von Hautknochen in der ventralen Wand des Abdomens. Bei Reptilien typisch, fehlen sie bei Vögeln, z. B.:
"Neben den echten Rippen zeigt das Exemplar des British Museum eine große Anzahl sogenannter ventraler Rippen oder Gastralia, Elemente von Hautknochen, die in der ventralen Wand des Abdomens liegen." (de Beer 1954, S. 18)
"Die Gastralia des Berliner Exemplars sind identisch mit denen des Exemplars des British Museum, aber mehr davon sind erhalten." (de Beer 1954, S. 19)
"Das „neue" Exemplar wurde am 8. September 1970 im Teyler Museum, Haarlem, Niederlande, ausgestellt gefunden. Es besteht aus zwei kleinen Platten (Exemplare 6928 & 6929), Teil und Gegenstück, die Abdrücke oder Teile der linken Hand und des Unterarms, des Beckens, beider Beine und Füße sowie einige Gastralia enthalten." (Ostrom 1970, S. 538)
"Außerdem sind zahlreiche Fragmente von Gastralia vorhanden, schwache Abdrücke von drei oder vier dorsalen Wirbeln, . . " (Ostrom 1972, S. 291).
"Das Gegenstück (Nr. 6929) enthält zusätzliche Gastralia, Phalangen, .." (Ostrom 1972, S. 291)
"Gastralia, oder Hautknochen-Rippen des Abdomens, sind in allen fünf Skelettexemplaren von _Archaeopteryx_ vorhanden" (Ostrom 1976, S. 139-140).
Gastralia sind am Eichstatter Exemplar vorhanden (Siehe Wellnhofer 1974, Abb. 7C)
Tabelle der Archaeopteryx-Merkmale
1 = vorhanden; * = vorhanden bei einigen; ? = möglicherweise vorhanden; x = nicht vorhanden
Dinosaurier Archae Vögel
1 * 1 1
2 x 1 1
3 * 1 1
4 * 1 1
5 x x 1
6 x x 1
7 * x 1
8 * 1 x
9 1 1 x
10 1 1 x
11 1 1 x
12 1 1 x
13 1 1 x
14 1 1 x
15 1 1 x
16 6 6 11-23
17 1 1 x
18 1 1 *
19 1 1 x
20 1 1 x
21 1 1 x
22 1 1 x
23 1 1 x
Es lässt sich erkennen, dass Archae viele mehr Merkmale aufweist, die in Dinosauriern und nicht in Vögeln vorkommen, als Merkmale, die in Vögeln, aber nicht in Dinosauriern vorhanden sind. Dies ist der Grund, warum Archae eine echte Übergangsart ist, da sie einige Merkmale teilt, die für eine Gruppe diagnostisch sind, während sie gleichzeitig Merkmale beibehält, die für ihre Stammesgruppe diagnostisch sind. Jeder, der behauptet, Archae sei zu 100 % Vogel, liegt falsch. Jeder, der behauptet, das Skelett von Archae sei sogar überwiegend vogelartig, liegt falsch. Jeder, der behauptet, Archae habe einen „vollständig vogelartigen" Schädel, liegt falsch.
Dieser letzte Punkt wird im Bezug auf die Behauptung von Dr. Duane Gish gemacht, dass das Schädelskelett von Archae "völlig vogelähnlich" sei (R. Trott, persönliche Mitteilung 1994). Diese Behauptung ist falsch. Um dies zu zeigen, müssen wir das Schädelskelett von Archae weiter betrachten.
Kraniale Merkmale von Archaeopteryx
Wie oben erwähnt, behauptet Dr. Gish, dass das Schädelskelett von Archae „völlig vogelartig" sei. Dies ist falsch. Romer (1950, S. 261) beschreibt Archae wie folgt: „Der Schädel ist, soweit ersichtlich, eher vogelartig. . . ". Allerdings ist dies weit entfernt von „völlig vogelartig", und Romer stützte sich dabei auf die detaillierte Rekonstruktion des Berliner Exemplars durch Heilmann (1926). Ostrom (1976, S. 131) äußert sich zur Heilmann-Rekonstruktion wie folgt:
"Trotz der dort gezeigten Details [Heilmanns Rekonstruktion des Schädels -cn] erlaubt das tatsächliche Exemplar keine derart detaillierten und präzisen Schlussfolgerungen. Der Schädel des Berliner Exemplars - cn - ist stark zerdrückt, und die Knochen sind ausgedehnt gebrochen, abgeplatzt und verzerrt - in einem Ausmaß, dass sehr wenige Schädel- oder Kiefernähte eindeutig identifizierbar sind. Heilmanns Rekonstruktionen wurden von vielen Autoren neu veröffentlicht, und darauf basierende Interpretationen und Hypothesen. Wahrscheinlich waren einige Autoren sich der unzureichenden Grundlage von Heilmanns Rekonstruktion nicht bewusst, und das ist verständlich, wenn sie keine Gelegenheit hatten, das Exemplar selbst zu untersuchen."
Und wieder auf derselben Seite:
"Glücklicherweise bietet das Eichstatter Exemplar nun eine vergleichende Grundlage zur Bewertung und Korrektur früherer Rekonstruktionen des Berliner Schädels."
Wie oben erwähnt, wurde das Eichstatter Exemplar erst 1974 beschrieben, daher basierte Romers Beschreibung auf der Rekonstruktion von Heilmann.
Mit beiden Exemplaren hat Ostrom (1976 S. 132) 10 Merkmale am Schädel von Archae beschrieben, die es mit anderen theropoden Dinosauriern wie Ornitholestes, Compsognathus, Velociraptor und möglicherweise Saurornithoides gemeinsam hat. Diese sind:
- Ein scharf zugespitzter Schnabel. Bei Vögeln besteht der „Schnabel" aus dem Maxillarknochen. Obwohl der Schnabel bei Archae verkürzt ist.
- Lange, elliptische äußere Nasenöffnungen [Nasenlöcher], die fast ausschließlich oben und unten von der Prämaxilla und dem Nasenbein (ein Knochen am vorderen Teil des Oberkiefers) begrenzt werden. Bei Vögeln sind die Nasenöffnungen aufgrund der Vergrößerung der Prämaxilla verkleinert und nach hinten (näher an den Augen) verschoben.
- Ein großes präorbitales Fossa und ein großes, dreieckiges postorbitales Fossa, das zwei kleine Öffnungen enthält. (Dies sind Löcher im Schädel. Eines ist eine einzelne Öffnung vor dem Orbit oder Augenhöhlen, das andere befindet sich hinter dem Augenhöhlen und hat zwei Öffnungen). Bei anderen fossilen Vögeln ist das präorbitale Fossa verkürzt, das postorbitale Fossa hat nur eine einzelne Öffnung. Bei modernen Vögeln ist das präorbitale Fossa stark verkürzt und das postorbitale Fossa hat eine einzelne Öffnung.
- Ein schlanker, nahezu vertikaler präorbitaler Balken, der das antorbitale Fossa und den Orbit trennt (Da ist wieder dieses Loch! Das bedeutet, dass es einen vertikalen Knochenbalken gibt, der das Loch vor dem Auge vom Auge selbst trennt). Fossile und moderne Vögel haben einen viel robusteren präorbitalen Balken – aufgrund der Verkürzung des präorbitalen Fossa.
- Ein großer, runder Orbit, der einen großen Sklerotischen Ring enthält. (Der Sklerotische Ring ist bei Reptilien, Vögeln und Actinopterygier-Fischen verbreitet, aber die meisten fossilen Reptilien und alle fossilen Vögel besitzen ihn. Es handelt sich um eine Reihe von verknöcherten Platten, die den Augapfel umgeben).
- Ein dünner, gerader Jugal-Knochen bildet den Zygoma-Bogen (dies ist der Knochen, der unter dem Auge verläuft, das Wangenknochen, wie es sich bei Menschen nach hinten zum Ohr erstreckt). Bei modernen Vögeln besteht der Bogen aus den Quadratojugal-, Jugal- und Maxillarknochen.
- Ein kräftiges, mäßig langes Quadrat, das nach vorne geneigt ist. Dies ist der Knochen im Oberkiefer, der Teil der Kiefergelenkverbindung – mit dem Articularknochen – bei Reptilien ist, und Archae hat ein großes, um, wenn Sie verstehen, was ich meine! (Übrigens – für diejenigen unter Ihnen, die noch dabei sind, ist das Quadrat am Steg im Oberkiefer befestigt, und wie wir alle wissen, ist der Steg der Knochen, der im Ohr vibriert, damit wir alle hören können. Somit waren Steg und Quadrat bei Reptilien verbunden, und es ist kein großer Sprung nach vorne, wenn sowohl Steg als auch Quadrat im Ohr vorhanden sind, wie es bei Säugetieren der Fall ist. Somit ist die Behauptung von Gish, dass – damit die Knochen beim Übergang von Reptilien zu Säugetieren in das Ohr gelangen könnten, sie durch eine Phase gehen müssten, in der sich das „Säugetier" jedes Mal taub machen würde, wenn es den Mund öffnete – nicht zutreffend ist, da der Zustand der Artikulation von Steg und Quadrat im Kiefergelenk bei fossilen und lebenden Reptilien gefunden wird, aber ich schweife ab).
- Ein Unterkiefer, der ungewöhnlich flach ist und hinter der Zahnreihe eine auffällige Biegung aufweist. Nicht bei Vögeln.
- Ein langer retroartikulärer Fortsatz. Wiederum ein klassisches Merkmal des Reptilien-Kiefergelenks, aber auch bei Vögeln geteilt.
- Prämaxilla-, Tränen- und Jugal-Knochen sind getrennt. Bei Vögeln ist der Prämaxillarknochen mit dem Jugal-Knochen verbunden; der Jugal-Knochen ist mit dem Tränenknochen verbunden. . .
:-)
- Einziger Vomer.
- Starke Krümmung des Ektopterygoide-Hakens.
- Ein dorsaler Ausschnitt im Prootic.
Obwohl Archaeopteryx bestimmte Schädelmerkmale aufweist, die avian sind, reichen diese nicht aus, um die Charakterisierung „vollständig vogelartig" zuzulassen.
Heilmann beschrieb eine externe mandibuläre Fenestra (ein Loch im Unterkiefer), die von unten durch das Dentaleum begrenzt wird – was der Zustand bei Vögeln ist – statt durch das Angulareum, wie es bei Reptilien der Fall ist. Dies würde darauf hindeuten, dass Archae einen Unterkiefer besitzt, der sowohl vogelartige (genannte mandibuläre Fenestra) als auch reptilienartige Merkmale (zahntragender Unterkiefer) aufweist. Das ist alles sehr gut und schön und ist ein Merkmal, das man bei Archae gerne sehen würde. Allerdings weist Ostrom (1976, S. 132) darauf hin:
"Gerade so sehr ich diese Interpretation gerne akzeptieren würde, so macht der hochgradig zersplitterte Zustand des Unterkieferknochens (oder der Knochen), die die vermeintliche mandibuläre Fenestra entweder von unten oder von oben begrenzen, es unmöglich, ihre Identifizierung zu bestätigen. Tatsächlich lassen die zersplitterten oberen Ränder der vermeintlichen Fenestra erhebliche Zweifel an der Existenz einer 'Fenestra' aufkommen – ein Zweifel, der durch das Eichstatter Exemplar nicht beseitigt wurde." So ist es denn keineswegs so, dass die Evolutionisten verschwören, um Archae in dem bestmöglichen Licht erscheinen zu lassen; eine sehr nützliche Eigenschaft, die bei der Beschreibung von Archae als echtes Übergangsfossil geholfen hätte, hat sich in der Tat höchstwahrscheinlich als nicht vorhanden erwiesen. So viel von der evolutionistischen Verschwörung! Tatsächlich zeigt das siebte Exemplar, dass eine mandibuläre Fenestra fehlt (Elzanowski & Wellnhofer 1996)
Ein weiteres wichtiges Merkmal des Schädels von Archae ist der Occipitalfortsatz und das Foramen magnum. Bei Archae liegen diese deutlich über dem dorsalen Ende des Quadrats. Wie Ostrom (1976, S. 136) schreibt:
"Dieser primitive Zustand ist charakteristisch für sowohl Pseudosuchia als auch Theropoden, im Gegensatz zu allen späteren Vögeln, bei denen der okzipitale Kondylus und das Foramen magnum an der Basis des Schädels liegen, deutlich unterhalb des Niveaus des oberen Extremitätsendes des Quadrats. In diesem Merkmal war Archaeopteryx weit entfernt von der Vogelähnlichkeit."Whetstone (1983, p. 449) does describe the Schädelkapsel (as opposed to the Schädel) of Archaeopteryx as, "typically avian", however, he also describes skull features found in Archaeopyteryx and not in birds.
In jüngerer Zeit gab es einige Spekulationen über die Struktur des Quadrats bei Archaeopteryx. Das Quadrat des Eichstätter Exemplars wurde als einköpfig beschrieben – d. h. die Oberseite des Quadrats besitzt nur einen abgerundeten Knopf, der sich an nur einer Stelle mit dem Schädel verbindet. Bei allen modernen Vögeln ist das Quadrat zweiköpfig – d. h. die Oberseite des Quadrats weist zwei abgerundete Knöpfe auf und verbindet sich daher an zwei Stellen mit dem Schädel. Basierend auf Computertomographie schlugen Haubiltz et al. (1988) vor, dass Archaeopteryx ein zweiköpfiges Quadrat besessen habe. Das Bild ist jedoch schlecht, und das Vorhandensein eines weiteren Knochens unter dem Quadrat kann nicht ausgeschlossen werden. Das einzige in situ gefundene Quadrat bei einem Exemplar ist einköpfig, und ein ähnlicher Knochen vom Londoner Exemplar, der als Quadrat erscheint, ist ebenfalls einköpfig. Die Identifizierung des Quadrats beim siebten Exemplar bestätigte, dass dieser Knochen einköpfig war (Elzanowski & Wellnhofer 1996). Somit erscheint das Quadrat von Archaeopteryx reptilisch und nicht vogelartig.
Wie zu sehen ist, ist Archae's Schädel nicht "vollständig vogelartig". Die Behauptung, er sei "vollständig vogelartig", ist unbegründet.
Einerseits kann die beobachtete Differenz zwischen _Archaeopteryx_, fossilen Vögeln und modernen Vögeln als bloße Variation innerhalb der Vögel erklärt werden. In diesem Zusammenhang steht die Behauptung, dass einige moderne Vögel (Ratiten) eine Reihe morphologischer Merkmale mit _Archaeopteryx_ teilen.
Archaeopteryx + moderne Vögel, nur Variation innerhalb einer Art?
Es wurde vorgeschlagen, dass die Unterschiede zwischen Archaeopteryx und modernen Vögeln eine einfache Variation innerhalb der Gruppe darstellen. Dies ist jedoch nicht korrekt. Moderne Vögel zeigen eine große Anzahl abgeleiteter morphologischer Merkmale, die Archaeopteryx nicht besitzen. Morphologisch erscheint Archaeopteryx eindeutig enger mit Theropoden-Dinosauriern verwandt als mit jeder anderen Gruppe und wird aufgrund des Besitzes von nur zwei Hauptmerkmalen, nämlich der Anwesenheit von Federn und der Anwesenheit eines vollständig zurückgebildeten Hallux (Zeh), mit Vögeln und nicht mit Theropoden-Dinosauriern gruppiert.
Die relevanten morphologischen Variationen können wie folgt (wenn auch grob) grafisch dargestellt werden:
|
^ | A = Archaeopteryx
| |
| ___________________
T | | |
i | | MODERNE VÖGEL |
m | __ __ __ __ __| |
e | | |___________________|
| | FOSSILE |
| __| VÖGEL |
| | A|__ __ __ __ __ __|
| |___|
|______________________________________________
"Reptil" "Vogel"
<--- Morphologie -->
Wie zu sehen ist, zeigt die Variation innerhalb der Vögel einen deutlichen Trend. Die eher "reptilienartigen" Morphologien treten bei den frühesten Vögeln auf, während die typische "moderne" Vogel-Morphologie auf spätere Vögel beschränkt ist. Wenn, wie angenommen, die morphologische Variation lediglich eine Variation innerhalb der Vögel darstellt, würden wir erwarten, dass die verschiedenen morphologischen Gruppen (fossile Vögel, moderne Vögel) gleichmäßig über den relevanten Zeitraum verteilt sind. Wenn jedoch Vögel von theropoden Dinosauriern abstammen, würden wir erwarten, dass die ersten Vögel mehr "reptilienartige" Merkmale aufweisen und die abgeleiteteren, modernen Vögel weniger "reptilienartige" Merkmale besitzen. Genau dies beobachten wir tatsächlich. Somit unterstützt die Verteilung der Merkmale innerhalb der Vögel deren Herleitung von theropoden Dinosaurier-Vorfahren und stützt nicht die Behauptung, dass die Variation lediglich 'innerhalb der Art' stattfindet.
Ratiten
Der morphologischen Merkmale der Ratiten (Strauße, Kiwis usw.) wird viel Bedeutung beigemessen, und oberflächlich betrachtet scheint es, als würden diese Vögel die These der 'Variation innerhalb der Art' stützen, wie die folgende Abbildung zeigt:
_____________________________
| | |
|RATITES| MODERNE VÖGEL |
|_______| |
|_____________________|
________________________________________________
"Reptil" "Vogel"
<--- Morphologie -->
Wichtig ist jedoch, dass andere morphologische Merkmale, die Ratiten besitzen, deutlich zeigen, dass sie von einer modernen, flugfähigen Vogelmorphologie abstammen und somit die Ratiten tatsächlich neotonisch gegenüber modernen Vögeln sind, wobei ihre eher „reptilienartigen“ Merkmale auf eine frühzeitige Beendigung der Entwicklung zurückzuführen sind, was ihnen einige morphologische Merkmale ähnlich wie bei Archaeopteryx und einigen fossilen Vögeln verbleibt (siehe hierzu). Diese Merkmale sind entwicklungsbiologischer Natur und liefern daher weitere stützende Belege für eine theropode Dinosaurier-Vogel-Verbindung.
Die Geschichte von zwei Becken
Dinosaurier können in zwei Gruppen eingeteilt werden, basierend auf der Form des Beckens. Die Saurischia, oder „Echsenbecken"-Dinosaurier, und die Ornithischia, oder „Vogelbecken"-Dinosaurier. Seltsam mag es erscheinen, dass Archaeopteryx und alle modernen Vögel sich von den Saurischia, nicht von den vogelbeckenartigen Ornithischia entwickelt haben sollen! Seltsam? Nun, eigentlich nicht. Die betreffenden Becken sind in Abb. 1 dargestellt.
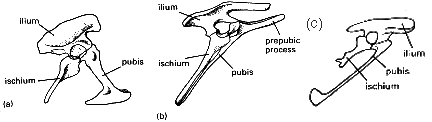
Das saurischische Becken (Abb. 1a) ist die ursprünglichere Struktur und weist den Schambeinfortsatz nach vorne und das Sitzbein nach hinten. Bei den ornithischischen Dinosauriern (Abb. 1b) verläuft der Schambeinfortsatz parallel zum Sitzbein nach hinten. Zusätzlich ist ein Vor-Schambeinfortsatz vorhanden, der als Ansatzpunkt für Muskeln angenommen wird, wenn das Schambein fehlt (wahrscheinlich zur Unterstützung des Darms). Diese Situation ist oberflächlich ähnlich der bei Vögeln (daher der Name „vogelbecken" – das Becken der Vögel war den Anatomen bereits bekannt, als die Dinosaurier beschrieben wurden). Bei genauerer Betrachtung zeigen sich jedoch deutliche Unterschiede. Bei Archaeopteryx (Abb. 1c) und allen Vögeln fehlt der Vor-Schambeinfortsatz und, im Gegensatz zu den saurischischen Dinosauriern, treffen sich die Schambeine nicht entlang der Mittellinie. Auch bei Archaeopteryx behält das Schambein die clubartige Endform bei, wie bei vielen saurischischen Dinosauriern. Es sollte auch beachtet werden, dass mehrere saurischische Dinosaurier ein nach hinten gerichtetes Schambein aufweisen, z. B. Troodon. Das Becken von Archaeopterys ist morphologisch intermediär zwischen saurischischen Dinosauriern und Vögeln.
Aber warum trat dies auf? Nun, der primäre Schubgenerator bei früheren Theropoden war der Caudofemoralis longus, der vom Bein bis zu einer gewissen Entfernung entlang des Schwanzes reichte. Spätere Formen wie die Dromaeosauriden (die Gruppe, die eng mit Archaeopteryx verwandt ist) ordneten dies neu an, sodass der primäre Schub durch Muskeln erzeugt wurde, die vom Bein zum Becken und zur Schwanzbasis reichten. Dies ermöglichte es, den Schwanz zu stützen und als stabilisierendes Gerät zu verwenden, wie bei Allosauriern und „Raptoren", oder ihn zu verlieren, wie bei modernen Vögeln.
Bei den Ornithischier wahrscheinlich trat der rückwärts gerichtete Pubis auf, um die Erweiterung von Magen und Darm zu ermöglichen, um pflanzliches Material besser verdauen zu können.
Die Untersuchung der embryonalen Entwicklung des modernen Vogelschädels ist ebenfalls lehrreich. Der embryonale Beckenbereich bei modernen Vögeln beginnt mit einer sehr ähnlichen Konfiguration wie bei Archaeopteryx, wobei das Ilium und das Os pubis in einem Winkel von ca. 45 Grad voneinander abgewinkelt sind. Die embryonalen Knochen drehen sich dann sekundär durch Winkel, die bei modernen Ratiten zu beobachten sind, bis sie die parallele Ausrichtung erreichen, die für erwachsene, moderne Vögel typisch ist (siehe Romanoff 1960, S. 1010):
Hühnchenembryo Winkel Ilium-Pubis Erwachsene mit gleichem Winkel
7.0 Tage 45 Grad Archaeopteryx
7.3 Tage 35 Grad Kiwi
8.0 Tage 30 Grad Strauß
Schlüpfen <5 Grad Modernes Huhn
Der Winkel zwischen dem Ilium und dem Os pubis im Becken moderner Ratiten liegt zwischen Archaeopteryx und modernen Vögeln, wie es zu erwarten wäre, wenn:
A) Moderne Vögel stammen von reptilienartigen Vorfahren ab, die denen von Archaeopteryx ähneln
B) Ratten sind neotene Nachkommen von fliegenden Vögeln, bei denen die Entwicklung vorzeitig gestoppt wurde.
Fliegen oder nicht fliegen
Fliegen ist ein schwieriges Geschäft. Allerdings gewährt Fliegen einen so starken evolutionären Vorteil, dass es nicht überraschend ist, dass die Fähigkeit sich mehrfach entwickelt hat. Der Erfolg der Vögel liefert ausreichende Belege für die positiven Vorteile des Fliegens. Wenn Archae geflogen wäre, hätte es wahrscheinlich viel die gleiche Methode wie heutige Vögel genutzt, daher muss eine Diskussion über die Möglichkeit des Fliegens bei Archae jene Strukturen berücksichtigen, die für das Fliegen bei Vögeln am relevantesten sind. Dies sind die Federn, die Flexibilität des Flügels, die Muskelmasse und die Anwesenheit eines keilförmigen Brustbeins.
Federn
Federn bestehen aus einem langen, sich verjüngenden Mittelstrang (Rachis), der dicht beieinander liegende Seitenäste trägt, die als Federvorsteher (Barben) bezeichnet werden. Die Barben auf beiden Seiten des Rachis bilden eine Fläche, die als Federblatt (Vane) bezeichnet wird. Die beiden Federblätter einer Feder können symmetrisch sein (d. h. dieselbe Breite aufweisen) oder asymmetrisch; in diesem Fall erscheint der Rachis einer Kante der Feder näher als der anderen. Die Flugfedern moderner Vögel sind in der Regel asymmetrisch, während Körperkontur- und Halbspaltfedern symmetrisch sind.
Bei modernen Vögeln sind die Remiges, oder Flügeldeckenfedern, stark für den kraftvollen Flug modifiziert (z. B. McFarland et al. 1985), hauptsächlich dadurch, dass der Schaft zur Vorderkante der Feder verschoben ist (d. h. die Vorderkante ist dünner als die Hinterkante), was zu einer asymmetrischen Feder führt. Die dünnere oder vordere Kante der Feder überlappt die breitere oder hintere Kante der Feder vor ihr (Abb. 2). Die hintere Kante enthält Zonen von Reibungsbarteln, die die überlappende Feder greifen und verhindern, dass die Federn zu weit auseinander gleiten.
Abb. 2
* * * O * * * * *
* * * O * * * * *
* * * O * * * * *
| | |
| | |--Steuerruder
| |---------Rachis
|-----------Vorderflügel
Speakman & Thomas (1994) vergleichen die Asymmetrie einiger Federn des Archaeopteryx (Flugfedern 4, 5 und 6) mit denen von modernen fliegenden und flugunfähigen Vögeln sowie der isolierten Feder aus Solnhofen. Sie stellten fest, dass die durchschnittliche Asymmetrie für Archaeopteryx-Federn 1,25 betrug, was niedriger war als bei modernen fliegenden Vögeln (niedrigste Werte um 2,2), jedoch mit dem Bereich moderner flugunfähiger Vögel überlappte. Die isolierte Feder zeigte eine Asymmetrie von 2,2 – genau im Bereich moderner fliegender Vögel. Es ist jedoch nicht bekannt, ob diese Feder von Archaeopteryx stammt oder, falls sie eine Archaeopteryx-Feder ist, wo auf Archaeopteryx sie sich befand. Norberg (1995) wies korrekt darauf hin, dass auch die Krümmung der Feder für die Flugfähigkeit wichtig ist, und die Federn des Archaeopteryx weisen eine signifikante Krümmung auf. Es scheint, als ob Speakman & Thomas möglicherweise die falschen Federn gemessen haben. Paul Davis (pers. komm. 1996) hat darauf hingewiesen, dass Speakman & Thomas die Primärfedern offenbar mit der Nummerierung am Flügelspitze beginnen und sich zum Körper hin bewegen, während die aktuelle Praxis darin besteht, die Federn mit der Nummerierung am Körper beginnen und sich nach außen entlang des Flügels zu bewegen (z. B. Rietschel 1985). Dies ist wichtig, da die Asymmetrie der Federn zur Flügelspitze hin abnimmt – somit scheinen Speakman & Thomas Federn mit geringerer Asymmetrie gemessen zu haben. Messungen der Federn mit höherer Asymmetrie in der Nähe des Körpers ergeben Werte von etwa 2,32 – was den unteren Bereich für existierende fliegende Vögel überlappt (z. B. Norberg 1995; P.G. Davis pers. komm. 1996). Die ersten Flugfedern von Vögeln sind jedoch immer hochgradig asymmetrisch, und so ist dieses Ergebnis mehrdeutig. Die meisten Flugfedern des Archaeopteryx scheinen eine etwas geringere Asymmetrie aufzuweisen als die bei heutigen fliegenden Vögeln gefunden. In der Realität jedoch ist die Federasymmetrie bei der Frage der Flugfähigkeit ein Rote-Indikator. Federn sind asymmetrisch, um die aerodynamische Integrität aufrechtzuerhalten, da sie sich unter aerodynamischen Lasten im Flug verformen, und somit tritt die größte Asymmetrie bei Vögeln auf, bei denen solche Lasten am größten sind – d. h. Vögel mit hoher Manövrierfähigkeit und Steigflug (J.M.V. Rayner, pers. komm. 1995). Somit ist Asymmetrie eine abgeleitete Bedingung für eine abgeleitete Flugfähigkeit – eine, die Archaeopteryx niemals besessen hat. Daher ist es nicht überraschend, dass Archaeopteryx Federn mit geringer Asymmetrie besaß, aber dies schließt Archaeopteryx nicht von Flug aus.
Flügelflexibilität
Betrachten Sie den Flügel der Carniaren [Vögel mit einem keilförmigen Brustbein]. Ein Wunder der Anpassung, das gut in der Lage ist, große Tiere bei energieverbrauchendem Flug über lange Strecken zu tragen. Aber was ist es an der Anatomie des Flügels, das die Möglichkeit des Fluges ermöglicht? Nun, ganz einfach ausgedrückt: „Es liegt alles in der Handgelenkaktion."
Bei modernen Vögeln wirkt das Schultergelenk als Universalgelenk mit einem hohen Maß an Beweglichkeit, nicht nur bei Adduktion [Flügelschlag nach unten], Abduktion [Flügelschlag nach oben] und Extension [Ausstrecken des Flügels vom Körper weg] sowie Retraction [Einziehen des Flügels in den Körper], sondern auch bei signifikanter Rotation entlang der Längsachse des Humerus.
Der Ellbogen ist hingegen ein sehr eingeschränktes Gelenk. Er erlaubt nur planare Extension und Flexion [mit anderen Worten: Aufklappen und Zuklappen des Flügels – d. h. es gibt keine rotatorische Komponente].
Jedoch ist es das Handgelenk, das von Bedeutung ist. Das Handgelenk besteht aus zwei knöchernen Handwurzelknochen, die die Bewegung des Gelenks steuern. Distal davon liegt der verschmolzene Handwurzel-Mittelhandknochen – entstanden durch die Verschmelzung der verbleibenden Handwurzel [Handgelenk]-Knochen und der Mittelhandknochen [unterer Handknochen]. Daran schließt sich die manchmal unvollständig verschmolzene Hand an, die aus drei Fingern besteht. Diese komplexe Struktur, distal des Handgelenks, bildet eine nahezu starre Plattform für die Befestigung der primären Remiges [Flugfedern]; die sekundären Remiges setzen sich am Unterarm fest.
Die Konfiguration des Handgelenks, das sich an zwei kleinen, runden Handwurzelknochen artikuliert, ermöglicht eine erstaunliche Bewegungsfreiheit. Um dies zu demonstrieren, müssen Sie bequem am Computerterminal sitzen. Strecken Sie Ihre Arme aus, als würden Sie gleich tippen (Handflächen nach unten, behaarte Stellen nach oben). Drehen Sie nun Ihre Hände so, dass die Handflächen nach oben zeigen (als würden Sie etwas tragen). Diese Drehung wird beim Vogel vom Handgelenk ausgeführt. Beachten Sie, dass beim Menschen diese Bewegung durch die Drehung des Unterarms und nicht des Handgelenks erfolgt. Bringen Sie nun die Hände wieder in die Position mit den behaarten Stellen nach oben und drehen Sie die Hände so, dass die Handfläche zum Computer zeigt (das universelle „Stopp"-Zeichen), und drehen Sie die Hand nach unten, bis die behaarten Stellen auf den Computer zeigen. Diese Drehung wird beim Vogel vom Handgelenk ausgeführt, wie auch beim Menschen.
Wie Sie nun gezeigt haben, ist das carniate Handgelenk viel beweglicher als das menschliche Handgelenk (Sie haben auch jedem Kollegen, der zuschaut, bewiesen, dass Sie verrückt sind und dringend Urlaub brauchen!). Diese Beweglichkeit ist für den motorisierten Flug von entscheidender Bedeutung, da sie es dem Handgelenk/-Hand ermöglicht, während eines vollständigen Schlages eine faule Acht zu beschreiben. Denken Sie daran, dass das Ellbogengelenk keine Rotation zulässt, aber der Flügel in der Lage sein muss, während des Abwärts- (Power-) Schlages eine feste, offene Flügelfläche zu präsentieren, während er während der Erholung (Aufwärts-) Schlag die Rotation des Flügels ermöglichen muss, um den Widerstand zu minimieren. Die Vögel erreichen dies, indem sie das Handgelenk und die Schulter rotieren.
An der Spitze des Auftriebsstrokes ist das Handgelenk so orientiert, dass es die maximale Oberfläche der Luft gegenüberstellt (Handfläche nach unten). Während des Auftriebsstrokes beschreibt Handgelenk und Hand eine „S"-Form oder zwei Bögen, die die linke Seite des oberen und die rechte Seite des unteren Loops einer „8" darstellen. Am Ende des Auftriebsstrokes befinden sich Handgelenk und Hand am unteren Punkt der „8". Zu Beginn des Rückholstrokes orientiert sich das Handgelenk um 90 Grad neu, wodurch die minimale Oberfläche der Luft gegenübergestellt wird. Der Flügel vervollständigt dann die „8", indem er die linke Seite des unteren und die rechte Seite des oberen Loops beschreibt. Es ist einfacher, dies auszuführen als zu lesen. Halten Sie Ihren rechten Arm waagerecht von der Schulter aus gestreckt, mit der Handfläche nach unten. Verriegeln Sie den Ellenbogen, sodass Sie den Arm nicht beugen, sondern nur rotieren können. Nun ist der Auftriebsstroke eine „S"-Form, wobei die Handfläche nach unten zeigt und der Ellenbogen verriegelt bleibt. Am unteren Punkt der „S" drehen Sie die Handfläche um 90 Grad, bis sie nach vorne zeigt (tatsächlich müssen Sie Ihren gesamten Arm drehen, um dies zu tun; bei Vögeln wird dies vom Handgelenk ausgeführt). Nun ist der Rückholstroke ein spiegelbildliches „S", das eine „8" vervollständigt; wenn Sie den oberen Punkt des Strokes erreichen, sollte die Handfläche zurückdrehen, um wieder nach unten zu zeigen, und Sie sind bereit für den Auftriebsstroke. Wiederholen Sie dies einige Male, bis Sie das Gefühl dafür bekommen oder bis jemand nach den Leuten in weißen Kitteln schickt. So können Vögel fliegen, und hoffentlich erkennen Sie nun die Bedeutung eines flexiblen Handgelenks.
Eine weitere wichtige Bemerkung zu diesem Abschnitt: Das Faltbewegen des Flügels erfolgt durch die Kontraktion des M. biceps brachii, der am Ulna befestigt ist. Diese Bewegung faltet automatisch den Metacarpus und die Hand nach unten und nach innen, sodass das Faltbewegen am Handgelenk den M. biceps brachii einbezieht.
Nun, der Punkt all dieses ist: Tritt diese Anordnung in der Archae auf? Tritt diese Anordnung in maniraptoren Theropoden auf? Falls ja, welchen möglichen Nutzen könnte dieser spezielle Handgelenk für maniraptoren Theropoden haben? Könnte dies ein Beispiel für Voranpassung sein (ach! Entschuldigung, dass ich das „P"-Wort benutze).
Zunächst zeigt das Handgelenk von Archae tatsächlich einige (aber nicht alle) Merkmale, die im modernen carnischen Handgelenk zu sehen sind. Am auffälligsten ist ein lunatkarpius, der halbkugelförmig ist, dessen flache Fläche mit dem Karpometakarpus articuliert und deren abgerundetes Ende mit dem Handgelenk articuliert. Dies bietet ein flexibles Handgelenk, obwohl nicht so flexibel wie bei modernen Carnivoren.
Zweitens wird diese Konfiguration in mehreren Theropoden-Taxa gefunden, wie Deinonychus, Velociraptor und Stenonychosaurus. Es gibt hier zwei Probleme: erstens postdatieren all diese Formen das Archae, sie sind alle kreidezeitliche Formen; zweitens, was wäre der Nutzen eines geschickten, flexiblen Handgelenks für eine Gruppe von Theropoden, die alle nur Zähne und Krallen sind?
OK, das erste Problem ist leicht zu lösen. Es handelte sich um ein Problem, das darin bestand, dass dieses Merkmal nur bei Cretazischen Theropoden und bei den späten Jurassischen Archae vorkam, da es, um ein wichtiges Merkmal zu sein und eine enge Verbindung zwischen Theropoden und Archae zu zeigen, Theropoden, die zeitgleich mit den Archae lebten, diese besondere Anordnung aufweisen mussten. Durch die Einführung von Coelurus fragilis, einem späten Jurassischen Theropoden aus Amerika, besitzt diese Form tatsächlich ein sehr ähnliches Handgelenk (Ostrom 1980), was nicht nur die Verbindung zwischen Theropoden und Vögeln, sondern auch die Verbindung zwischen Theropoden, Archae und Vögeln stärkt.
Das zweite Problem ist etwas schwieriger. Warum sollte ein solches Handgelenk bei maniraptoranen Theropoden erhalten bleiben, die es bei der Tötung eher auf Zähne und Krallen als auf das Handgelenk ankommen lassen? Ostrom (1995) schlägt vor, dass die Flexibilität dort nützlich wäre, wo eine Greifaktion der Hände die tödliche Wirkung der Zähne und/oder Krallen verstärken könnte. Ich würde noch weiter spekulieren. Bei modernen Vögeln sind die Bizeps an der Faltung des Flügels beteiligt. Wenn dies auf die Theropoden übertragen würde, würde die Bewegung, das Beute in den Körper zu ziehen, den Griff des Theropoden verstärken, indem das größte Muskel des Arms – der Bizeps – zum Einsatz kommt, anstatt nur die Handgelenksmuskeln. Ein solches Szenario würde einen deutlichen Vorteil bieten, und daher ist die Erhaltung des Handgelenks bei den Theropoden nicht überraschend. Der Titel des Sequels zu „Jurassic Park" ist somit leicht: „Jurassic Park II: Das Kuscheln der Coelosaurier“!
Es scheint also, dass das flexible Handgelenk zunächst bei Theropoden-Dinosauriern als Hilfe zum Ergreifen und Festhalten von Beute entstanden sein könnte und sekundär von den Vögeln als Mechanismus übernommen wurde, der den Flug ermöglichte. Es ist sehr wohl möglich, dass nicht nur die Theropoden-Dinosaurier die Vorfahren der Vögel waren, sondern dass sie die einzige Gruppe sein könnten, die die Ahne der modernen Vögel ist.
Jedoch scheint das Handgelenk von Archae nicht die Flexibilität aufzuweisen, um einen kraftvollen Flug zu ermöglichen.
Schultergelenk
Der Schultergelenk ist auch für moderne Vögel wichtig, da es nicht nur eine vollständige Adduktion (Flügelbewegung nach unten - Abb. 3b) ermöglicht, sondern auch eine vollständige Abduktion (Flügelbewegung nach oben - Abb. 3d)
Abb. 3 (von vorne betrachtet)
Adduktion Abduktion
a) -----( )----- c) -----( )-----
b) /( )\ d) | |
| | | |
| | \( )/
d. h. der Flügel kann über den Rücken gefaltet werden (versuchen Sie es und sehen Sie, wie schwierig das ist). Diese vollständige Bewegungsbreite ist wichtig, um modernen Vögeln das powered Fliegen zu ermöglichen. Es stellt sich heraus, dass das Schultergelenk von Archae eine Zwischenstellung in der Ausrichtung zwischen einigen Coelurosauriern (Theropoden-Dinosauriern, z. B. Deinonychus) und Vögeln (Jenkins 1993) einnimmt – genau wie man es von einer Übergangsform erwarten würde. Allerdings ermöglicht das Schultergelenk zwar einen erheblichen Grad an Arm (Flügel)-Hebung, reicht jedoch nicht aus, um eine vollständige Abduktion wie bei modernen Vögeln zu erlauben. Daher scheint das Schultergelenk von Archae die vollständige Bewegungsbreite nicht zu erlauben, die für das powered Fliegen benötigt wird.
Muskelmasse
Bei modernen Vögeln machen die Flugmuskeln (der Pectoralis) bis zu 35% des Gewichts des Tieres aus (denken Sie an all dieses saftige Fleisch am vorderen Ende eines Brathuhns oder einer Truthahn). Diese massive Muskelmasse liefert die für das Flügelschlag-Flug benötigte Leistung und die Fähigkeit, vom Stehen aus abzuheben. Ruben (1991) verwendete eine Schätzung der Pectoralis-Muskelmasse für Archae von ca. 9% des Körpergewichts. Dies ist viel niedriger als bei modernen Vögeln, ging aber davon aus, dass der Muskel an die knorpelige Brustwand und nicht an einen Kiel angeheftet war – da ein Kiel bei Archae unbekannt ist. Der Kiel ist eine knöcherne Verlängerung des Brustbeins für die Befestigung von überschüssiger Flugmuskulatur bei modernen Vögeln. Wenn die Muskeln bei Archae an die Brustwand angeheftet waren, wären die Pectoralis-Muskeln zu kurz gewesen, um eine vollständige Abduktion zu ermöglichen (siehe „Schultergelenk" oben). Allerdings ist die Anwesenheit eines kleinen, knorpeligen (und daher nicht erhaltenen) Kiels nicht ausgeschlossen. (Exemplare von Archie aus dem Solnhofener Kalkstein zeigen keine Hinweise auf ein Brustbein. Allerdings deuten jüngste Funde von Archie-ähnlichen Formen aus China darauf hin, dass Archae tatsächlich ein Brustbein besaß (J.M.V. Rayner, University of Bristol, pers.comm.).) Daher ist die 9%-Zahl ein Mindestwert. Dieser Wert liegt weit unter dem für moderne Vögel. Rubin schlug vor, dass die maximale Leistung der Lokomotionsmuskeln bei extanten aktiven Reptilien das Doppelte derjenigen von Vögeln und Säugetieren beträgt, was darauf hindeutet, dass, wenn Archae eine reptilische Physiologie besaß, ein Flügelschlag-Flug möglich war, selbst mit einer so kleinen Muskelmasse. Speakman (1993) stellte dies in Frage und zeigte, dass die von Rubin für Reptilien vorgeschlagene Leistung übertrieben war und dass Archae nicht über die Leistung verfügt hätte, um einen Flügelschlag-Flug aufrechtzuerhalten. Auch Speakman wies darauf hin, dass die theoretische untere Grenze der Pectoralis-Muskelmasse, die für das Abheben vom Stehen aus erforderlich ist, ca. 16% des Körpergewichts beträgt. Somit war Archae nicht in der Lage, einen Flügelschlag-Flug oder vom Stehen aus abzuheben.
Der Supracoracoideus ist ein wichtiger Muskel bei modernen Vögeln, da dieser Muskel den Flügel während des Fluges hebt oder abduziert. Archae besaß nicht die beim modernen Vogel vorkommende Anordnung des Supracoracoideus (Ostrom 1974). Daher scheint Archae weder über die Muskelmasse noch über die Muskelanordnung verfügt zu haben, um einen kraftvollen Flug zu ermöglichen oder von einer stehenden Startposition abzuheben.
Es scheint, dass die Struktur und Physiologie von Archae gegen den powered flight sprechen. Doch ist dies das Todesurteil für die akrobatischen Fähigkeiten von Archae? Tatsächlich nein. Beachten Sie die Verwendung des Ausdrucks "power flight" in der vorangegangenen Diskussion. Fast alle modernen fliegenden Vögel sind in der Lage, powered flight zu betreiben. Power flight ist kontrollierter, flatternder Flug bei niedrigen Geschwindigkeiten und die Fähigkeit, vom Stehen aus abzuheben. Diese Art des Fliegens erfordert eine große Muskelmasse und ist energetisch sehr aufwendig. Daher haben moderne Vögel eine Pectoralis-Muskelmasse, die etwa 20-35% der Körpermasse ausmacht. Um diese zusätzliche Masse aufzunehmen, hat sich auf dem Sternum ein Kiel entwickelt. In anderen evolutionären Entwicklungen sind die Flugfedern asymmetrischer geworden, um den powered flight zu unterstützen, und das Supracoracoideus-Muskelsystem hat die Funktion übernommen, das Heben oder Abduzieren des Flügels zu unterstützen, da dies bei niedrigen Fluggeschwindigkeiten schwierig ist.
Der powered flight ist zum Fliegen, was die 4-Minuten-Meile zum Laufen ist. Allerdings bedeutet es genau so wenig, dass jemand nicht in der Lage ist, eine 4-Minuten-Meile zu laufen, dass man daraus schließen kann, Archae sei nicht in der Lage zum Fliegen gewesen. Die für den Flug charakteristischen Merkmale, die wir bei modernen Vögeln sehen, sind abgeleitet, eine Anpassung an den powered flight, und ihr Fehlen (oder ihre Unterentwicklung) bei Archae kann daher nicht verwendet werden, um eine Unfähigkeit zum Fliegen zu suggerieren.
Die Struktur des Flügels von Archae deutet darauf hin, dass es nicht in der Lage zu einem kraftvollen, flatternden Flug bei niedrigen Geschwindigkeiten war. Allerdings deutet die Menge an Adduktion und Abduktion sowie die erhöhte Flexibilität des Handgelenks im Vergleich zu theropoden Dinosauriern darauf hin, dass eine Art von Flattern möglich war, ausreichend, um Archae in der Luft zu halten bei normalen Reisegeschwindigkeiten (ca. 7-9 ms^-1). Obwohl die geringe Masse des Pectoralis-Muskels einen kraftvollen Flug und das Starten vom Stehen auszuschließen scheint, sei darauf hingewiesen, dass einige fliegende Vögel eine Muskelmasse besitzen, die unter dem 16%-igen unteren Gewichtslimit liegt. Dazu gehören einige Ruderer (Grebes), die den Mangel an Muskelmasse durch eine lange laufende Startphase kompensieren (Speakman 1993). Archae ist gut mit einem terrestrischen Fortbewegungsapparat ausgestattet. Somit ist es möglich, dass Archae dieses Problem auf dieselbe Weise umgehen konnte wie die Ruderer.
Das Fliegen mit normalen Reisegeschwindigkeiten erfordert viel weniger Energie als der Kraftflug und somit weniger Muskelmasse, um ihn aufrechtzuerhalten. Als zusätzlicher Bonus ist das Flügel bei normalen Reisegeschwindigkeiten durch aerodynamische Kräfte im Aufschlag angehoben (abduziert) (Rayner 1988) – wodurch die Notwendigkeit der für den kraftvollen, flatternden Flug notwendigen Suprakorakoideus-Anordnung umgangen wird.
Archaeopteryx's Abstammung
Neben der theropodenartigen Abstammung wurden drei weitere vorgeschlagene Vorfahrengruppen für die Archae, die Ornithopoda, die Pseudosuchia und die Sphenosuchidae, genannt.
Die Theorie der Ornithopod-Vorfahren basierte auf dem Fuß und dem Becken bei verschiedenen Dinosauriern, die supposedly die bei Vögeln gefundene Bedingung annähern sollten. Allerdings sind die meisten der in der ursprünglichen Theorie von 1883-4 verwendeten Taxa nun als saurischisch und nicht als ornithischisch bekannt. Zwar gab es bei einigen Ornithischiern eine Tendenz zur Reduktion des Hallux (Zehengrundglied), doch war er nie opponierbar wie bei Archae. Es wurde vorgeschlagen, dass Archae ein sehr ornithischisches Becken besaß, jedoch hat jüngere Arbeit gezeigt, dass dies ein Artefakt der postmortalen Bewegung von Elementen der Beckenregion ist und es nur sehr wenig Ähnlichkeit mit dem ornithischischen Becken gibt. Mit diesem Befund verschwindet der Zusammenhang zwischen Ornithischiern und Archae, und es wird nun angenommen, dass kein Zusammenhang zwischen ihnen besteht.
Es wurde vorgeschlagen, dass Vögel aus einer fast unbekannten Reptiliengruppe des mittleren oder späten Trias, den Sphenosuchidae (Walker 1972, 1974), hervorgegangen sind, die auch zu den Krokodilen geführt haben. Die Theorie basiert auf einem einzigen Exemplar von Sphenosuchus. Allerdings sind weder der Schädel und die Kiefer, noch das Schulterblatt- und Schlüsselbein von Sphenosuchus in ihrer allgemeinen Konfiguration auch nur entfernt auf eine Verwandtschaft mit Archae (Ostrom 1976) hinweisend. Kürzlich hat der Hauptvertreter der Krokodil-Vogel-Hypothese erklärt, dass die Hypothese „so schwach geworden ist, dass sie sehr schwer zu stützen ist." (Walker 1985, S. 133)
Der Pseudosuchia-Verwandtschaftszusammenhang wurde zu Beginn dieses Jahrhunderts sehr stark befürwortet. Dies basierte jedoch weitgehend auf Heilmanns Rekonstruktion des Berliner Exemplars, die sich als ungenau erwiesen hat. Nach Ostrom (1976, S. 159):
"Es gibt sehr wenige anatomische Ähnlichkeiten zwischen Archaeopteryx und irgendeinem Pseudosuchier. Tatsächlich ist nur in einer Eigenschaft irgendein Pseudosuchier dem Archaeopteryx ähnlicher als irgendein Theropode, nämlich das Verhältnis von Tibia zu Femur bei Scleromochlus, Lagosuchus und Lagerpeton, wobei die Tibia um 20% bis 30% länger ist als das Femur. Unter den Theropoden ist die Tibia nur bei Struthiomimiden, Compsognathus, Microvenator und Deinonychus länger als das Femur, jedoch nur um 10% bis 15%. Bei allen anderen Merkmalen findet sich die engste Ähnlichkeit mit der in Archaeopteryx erhaltenen Morphologie bei coelurosaurischen Theropoden." [ursprüngliche Hervorhebung]
Chiappe (1995) liefert eine sehr gute Übersicht über die frühe Evolution der Vögel.
Protoavis
Einige Leute behaupten gerne, dass die Entdeckung eines fossilen Vogels aus dem Trias von Texas (Protavis) beweist, dass Archae nicht als Übergangsform zwischen Dinosauriern und Vögeln in Frage kommen kann, weil Protoavis Archae um 75 Millionen Jahre vorausgeht. Dies ist natürlich irriges Unsinn, hauptsächlich weil niemand behauptet, dass Archae die Übergangsart zwischen Dinosauriern und Vögeln sei, sondern lediglich, dass Archae eine Entwicklungsstufe darstellt, die die vorgeschlagene Abstammungslinie durchlaufen hat, um von Dinosauriern zu Vögeln zu gelangen. Archae ist, leid tut es mir zu sagen, evolutionär betrachtet, auf einem Ast. Es stellt einen Seitenzweig dar, der für vergleichende Zwecke nützlich ist, aber nicht im Zentrum der Dinge steht. Also selbst wenn es im Trias Vögel gegeben hätte, würde dieser Umstand die Bedeutung von Archae als Indikator dafür nicht schmälern, dass „ja, Vögel sich von Dinosauriern entwickelt haben könnten."
Allerdings beachten Sie das „wenn" im vorherigen Satz. Es gibt erhebliche Probleme mit Protoavis. Nach der Interpretation von Chatterjee (1991) sagt Ostrom (1991) dazu [meine Paraphrase]: Das einzige veröffentlichte Material aus dem Fossil ist eine Monografie in den Philosophical Transactions of the Royal Society of London. Allerdings beschreibt dies nur den Kopf. Dieser ist stark zerdrückt, und alle Teile wurden aus der Matrix extrahiert, was eine präzise Platzierung der Teile in Frage stellt. Die Beschreibung erfolgt aus einer avianen Perspektive, ohne Gegenperspektive (z. B. ist dies ein Dinosaurier?). Der Schädel ist so stark zerdrückt, dass diagnostische Merkmale nicht erhalten sind. Daher unterstützt das veröffentlichte Material die Ansicht nicht, dass dies ein Vogel ist. Tatsächlich zeigte eine Betrachtung des Fossils durch Ostrom (in freilich weniger als idealen Verhältnissen), dass die diagnostischen Merkmale, die das Fossil auf jede Weise identifizieren könnten, stark zerdrückt sind und es zweifelhaft ist, ob eine definitive Aussage durch das Fossil gestützt werden kann. Es kann ein Vogel sein, es kann es auch nicht sein.
Bitte beachten Sie, dass diese Infragestellung von Protoavis als Vogel kein „es kann kein Vogel sein, weil es vor Archae" evolutionistischer Gegenangriff ist. Wie bereits hervorgehoben wurde, schadet es selbst, wenn es ein Vogel ist, nicht der evolutionären Bedeutung von Archae.
Schlussfolgerungen
Archaeopteryx ist ein Vogel, weil es Federn besaß. Es behielt jedoch viele dinosaurische Merkmale, die bei modernen Vögeln nicht vorkommen, während es bestimmte Merkmale aufwies, die bei Vögeln, aber nicht bei Dinosauriern zu finden sind. Aufgrund dieser Tatsache stellt Archaeopteryx ein Beispiel für eine Gruppe in Übergang dar – einen Vertreter, der zwar am Rande des Übergangs von Dinosauriern zu Vögeln steht, ein Echo des tatsächlichen Ereignisses, trotzdem einen kurzen Einblick in den möglichen Mechanismus ermöglicht, der die Evolution der Vögel bewirkt hat, und durch seine bloße Existenz zeigt, dass ein solcher Übergang möglich ist.
Danksagungen
Dieses Ergebnis ist die Folge einer Reihe von Diskussionen mit Rich Trott, der sich gegen den systematischen Missbrauch von Archae durch bestimmte Gegner der Evolution wandte und dennoch Zeit fand, viele Verbesserungen für diesen Beitrag vorzuschlagen. Die Beiträge von Tom Holtz und Paul Davis werden ebenfalls anerkannt. Der Großteil der ursprünglichen Version wurde von Brett Vickers in HTML-Format für das TalkOrigins-Archiv konvertiert.
Dies ist eine paläontologische Veröffentlichung der University of Ediacara.
[Zurück zu den Archaeopteryx-FAQs]
Referenzen
Abbott, A. (1992) Archaeopteryx-Fossil verschwindet aus privater Sammlung. Nature, 357: 6.
Alexander, R.M. (1990) Tiere. Cambridge University Press, Cambridge. 509 S.
Barsbold, R. et al. (1990) Oviraptorosauria. In The Dinosauria, Weishampel, Dodson & Osmolska (Hrsg.) S. 249-258.
Barthel, K.W.; Swinburne, N.H.M. & Conway Morris, S. (1990) Solnhofen. Eine Studie zur mesozoischen Paläontologie. Cambridge University Press, Cambridge. 236 S.
Bellairs, A.D'A. & Jenkin, C.R. (1960) Das Skelett der Vögel. S. 241-300. In: Biologie und vergleichende Physiologie der Vögel, Band I, A.J. Marshal (Hrsg.). Academic Press, New York.
Britt, B.B.; Makovicky, P.J.; Gauthier, J. & Bonde, N. (1998) Postkraniale Pneumatisierung in Archaeopteryx. Nature, 395: 374-376.
Brooks, B. (1993) Pneumatische postkraniale Knochen bei Dinosauriern und anderen Archosauriern. Ph.D.-Dissertation, Department of Geology and Geophysics, University of Calgary, Calgary, Alberta, Kanada.
Bryant, H.N. & Russell, A.P. (1993) Das Vorkommen von Schlüsselbeinen innerhalb der Dinosaurier: Implikationen für die Homologie der avianen Furcula und die Nützlichkeit negativer Evidenz. Journal of Vertebrate Paleontology, 13(2):171-184.
Chatterjee, S. (1991) Schädelanatomie und Verwandtschaftsverhältnisse eines neuen triassischen Vogels aus Texas. Phil. Trans. R. Soc. Lond. B. 332: 277-342.
Chen Pei-Ji; Dong Zhi-Ming & Zhen Shuo-nan. (1998) Ein außergewöhnlich gut erhaltener Theropoden-Dinosaurier aus der Yixian-Formation Chinas. Nature, 391: 147-152.
Chiappe, L.M. (1995) Die ersten 85 Millionen Jahre der Vogel-Evolution. Nature, 378: 349-355.
Chure, D.J. & Madsen, J.H. (1996) Über das Vorkommen von Furkulae bei einigen nicht-maniraptorischen Theropoden. Journal of Vertebrate Paleontology, 16: 573-577. Colbert, E. H. & Morales, M. (1991) Evolution der Wirbeltiere: eine Geschichte der Wirbeltiere durch die Zeit. 4. Aufl. Wiley-Liss, New York. 470 S.
de Beer, G. A. (1954) Archaeopteryx lithographica. British Museum of Natural History, London.
de Beer, G.A. (1956) Evolution der Ratten. Bulletin of the British Museum (Natural History): Zoologie, 4: 57-70.
Elzanowski, A. & Wellnhofer, P. (1996) Schädelmorphologie von Archaeopteryx: Belege aus dem siebten Skelett. Journal of Vertebrate Paleontology 16 (1): 81-94
Gingerich, P.D. (1975) Evolutionäre Bedeutung der mesozoischen Zahnvögel. Smithsonian Contributions to Paleobiology, 27: 23-34.
Haubiltz, B.; Prokop, M.; Dohring, W.; Ostrom, J.H. & Wellnhofer, P. (1988) Computertomographie von Archaeopteryx. Paleobiology, 14: 206- 213.
Heilmann, G. (1926) Der Ursprung der Vögel. Witherby, London. 208 S.
Jenkins, F.A. (1993) Die Evolution des Schultergelenks bei Vögeln. American Journal of Science, 293A: 253-267.
Lambrecht, K. (1933) Handbuch der Palaeornithologie. Berlin. 1024 pp.
McFarland, W.N.; Pough, H.F.; Cade, H.F. & Heiser, J.B. (1985) Vertebrate Life, 2. Aufl. Macmillan Publishing, New York. 636 S.
McGowan, C. (1984) Am Anfang: Ein Wissenschaftler zeigt, warum die Kreationisten falsch liegen. Prometheus, Buffalo. 208 S.
Norberg, R.A. (1995) Federaussymmetrie in Archaeopteryx. Nature 374: 221.
Orr, R.T. (1966) Vertebratenbiologie, 2. Aufl. W.B. Saunders, Philadelphia. 483 S.
Ostrom, J.H. (1970) Archaeopteryx: Mitteilung über ein neues Exemplar. Science, 170: 537-538.
Ostrom, J.H. (1972) Beschreibung des Archaeopteryx-Exemplars im Teyler-Museum, Haarlem. Proceedings der Königlichen Niederländischen Akademie der Wissenschaften, B, 75: 289-305.
Ostrom, J.H. (1974) Archaeopteryx und der Ursprung des Fluges. The Quarterly Review of Biology, 49(1)
Ostrom, J.H. (1976) Archaeopteryx und der Ursprung der Vögel. Biological Journal of the Linnean society, 8(2): 91-182.
Ostrom, J.H. (1980) Coelurus und Ornitholestes: Sind sie dasselbe? In L. Jacobs (Hrsg.) Aspekte der Wirbeltiergeschichte. 245-256.
Ostrom, J.H. (1991) Vogel im Busch. Nature, 353: 212
Ostrom, J.H. (1995) Biometrie der Flügel und der Ursprung des Vogelzugs. Neues Jahrbuch für Geologie und Paläontologie, 195: 253-266.
Padian, K. (1998) Wann ist ein Vogel kein Vogel? Nature, 393: 729-730.
Qiang Ji; Currie, P.J.; Norell, M.A. & Shu-an ji. (1998) Zwei gefiederte Dinosaurier aus Nordostchina. Nature, 393: 753-761.
Rayner, J.M.V. (1988) Die Evolution des Vogel- und Säugetierflugs. Biological Journal of the Linnean Society, 34: 269-287
Rietschel, S. (1985) In: The Beginnings of Birds, Hecht, M.K., Ostrom, J.H., Viohl, G. & Wellnhofer, P. (eds.). pp. 303-318. Freunde Des Jura-Museums, Eichstatt.
Romanoff, A.L. (1960) Der Vogel-Embryo. Macmillan, New York. 1304 S.
Romer, A.S. (1950) Vertebratenpaläontologie. S. 261. University of Chicago Press, Chicago.
Ruben, J. (1991) Reptilienphysiologie und die Flugfähigkeit von Archaeopteryx. Evolution, 45: 1-17.
Speakman, J. R. (1993) Flugfähigkeiten in Archaeopteryx. Evolution, 47: 336-340.
Speakman, J.R. & Thomson, S.C. (1994) Flugfähigkeiten von Achaeopteryx. Nature, 370:514.
Tucker, B.W. (1938a) Funktionale evolutionäre Morphologie: der Ursprung der Vögel. S. 321-336. In: Evolution: Essays zu Aspekten der Evolutionsbiologie, G.A. de Beer (Hrsg.). Clarandon Press, Oxford.
Tucker, B.W. (1938b) Einige Beobachtungen zur Theorie von Dr. Lowe über die Beziehung der Struthioines zu den Dinosauriern und zu anderen Vögeln. Proceedings of the 8th International Ornithological Congress, Oxford. 1934: 222-224.
Unwin, D M (1998) Federn, Filamente und Theropoden-Dinosaurier, Nature 391: 119-120.
Walker, A. D. (1972) Neues Licht über die Entstehung von Vögeln und Krokodilen. Nature, 237: 257-263.
Walker, A. D. (1974) Evolution, organic. In Yearbook of Science and Technology, D.N. Lapedes (ed). pp. 177-179. McGraw-Hill, New York.
Walker, A. D. (1985) Der Schädelfundus von Archaeopteryx. In The Beginnings of Birds, Hecht, M.K., Ostrom, J.H., Viohl, G. & Wellnhofer, P. (Hrsg.). S. 123-134. Freunde Des Jura-Museums, Eichstatt.
Wellnhofer, P. (1974) Das fünfte Skelettexemplar von Archaeopteryx. Palaeontographica A, 147: 169-216.
Wellnhofer, P. (1993) Das siebte Exemplar von Archaeopteryx aus dem Solnhofener Kalkstein. Archaeopteryx, 11: 1-47.
Whetstone, K. N. (1983) Schädelkapsel von Mesozoischen Vögeln: I. Neue Präparation des "Londoner" Archaeopteryx. Journal of Vertebrate Paleontology, 2: 439-452.
Witmer, L. M. (1990) Das kraniofaziale Luftsack-System mesozoischer Vögel (Aves). Zoological Journal of the Linnean Society, 100: 327-378.
[Zurück zu den Archaeopteryx-FAQs]