|
por Raymond Sutera Copyright © 2001 [publicado: 10 de agosto de 2001] Este artículo apareció originalmente en Reports of the National Center for Science Education, una publicación de The National Center for Science Education. |
 |
|
¿Cómo convence alguien de que un fósil es un fósil transicional? ¿Rendirse? Es una pregunta trampa. No se puede. No hay forma de convencer a alguien que ya tiene una opinión formada. Pero a veces, es incluso peor. A veces, cuando señalas un fósil que cae en el centro de una brecha y es un extraordinario intermedio morfológico y cronológico, te responden: “Bueno, ¡ahora tienes dos brechas donde antes tenías solo una! ¡Estás retrocediendo!”
Uno de los desafíos más conocidos contra la existencia de fósiles transicionales entre los evolucionistas creacionistas es la supuesta falta de formas transicionales en la evolución de las ballenas. Duane Gish, del Institute for Creation Research (ICR), suele presentar repetidamente la “transición de joroba a espiráculo” para ridiculizar la idea de que las ballenas podrían haber evolucionado de ancestros terrestres con pezuñas. Simplemente no hay formas transicionales en el registro fósil entre los mamíferos marinos y sus supuestos ancestros mamíferos terrestres... Es bastante entretenido, partiendo de vacas, cerdos o búfalos, intentar visualizar cómo podrían haber sido los intermedios. Partiendo de una vaca, incluso podríamos imaginar una línea de descendencia que se extinguió prematuramente, debido a lo que podría llamarse un fallo mamario (Gish 1985: 78-9). Por supuesto, durante muchos años el registro fósil de las ballenas era bastante incompleto, pero ahora existen numerosas formas transicionales que ilustran la trayectoria de la evolución de las ballenas. Los descubrimientos recientes de fósiles de ballenas proporcionan la evidencia que convencerá a un escéptico honesto. Sin embargo, la biología evolutiva predice no solo la existencia de ancestros fósiles con ciertas características; también predice que todas las demás disciplinas biológicas deberían revelar también patrones de similitud entre las ballenas, sus ancestros y otros mamíferos correlacionados con la relación de evolución entre grupos. No debe sorprender que esto es lo que encontramos, y dado que los hallazgos en una disciplina biológica, como la bioquímica, se derivan sin referencia a otra disciplina, como la anatomía comparada, los científicos consideran que estos campos aportan evidencia independiente de la evolución de las ballenas. Como cabría esperar, estas líneas de evidencia independientes confirman todas el patrón de la evolución de las ballenas que anticiparíamos en el registro fósil. Para ilustrar este enfoque, presentaré la evidencia de múltiples campos para el origen de las ballenas a partir de mamíferos terrestres. Este texto examinará evidencia mutuamente reforzada de nueve áreas de investigación independientes. Por supuesto, como punto de partida, necesitamos describir lo que hace que una ballena sea una ballena. ¿Qué es una ballena? Una ballena es ante todo un mamífero, un vertebrado de sangre caliente que usa su alto metabolismo para generar calor y regular su temperatura interna. Las hembras de ballena dan a luz crías vivas, que amamantan con glándulas mamarias. Aunque las ballenas adultas no tienen cobertura de vello corporal, lo adquieren temporalmente como fetos, y algunas ballenas adultas tienen cerdas sensoriales alrededor de la boca. Estas características son inequívocamente mamíferas. Pero una ballena es un mamífero muy especializado con muchos caracteres únicos que no se comparten con otros mamíferos, y muchos de estos ni siquiera se comparten con otros mamíferos marinos como los sirénidos (manatíes y dugongos) y pinnípedos (focas, leones marinos y morsas). Por ejemplo, las ballenas tienen cuerpos hidrodinámicos que son gruesos y redondeados, a diferencia de los cuerpos generalmente delgados y alargados de los peces. La cola de una ballena tiene aletas horizontales, que son su único medio de propulsión a través del agua. La aleta dorsal está reforzada por tejido conectivo, pero es carnosa y totalmente sin huesos de soporte. Las vértebras cervicales de la ballena están acortadas y al menos parcialmente fusionadas en una sola masa ósea. Las vértebras detrás del cuello son numerosas y muy similares entre sí; los procesos óseos que conectan las vértebras están muy reducidos, lo que permite que la espalda sea muy flexible y produzca fuertes impulsos con las aletas de la cola. Las aletas que permiten a la ballena girar están compuestas por huesos del brazo aplanados y acortados, huesos de la muñeca planos en forma de disco y dedos múltiples alargados. La articulación del codo es prácticamente inmóvil, haciendo la aleta rígida. En la cintura escapular, la escápula está aplanada y no hay clavícula. Algunas especies de ballenas aún poseen una pelvis vestigial, y algunas tienen extremidades posteriores muy reducidas y no funcionales. La caja torácica es muy móvil: en algunas especies, las costillas están totalmente separadas de la columna vertebral, lo que permite que el tórax se expanda mucho cuando la ballena respira y que el tórax se comprima en profundidad cuando la ballena se sumerge profundamente. El cráneo también tiene un conjunto de características únicas entre los mamíferos. Las mandíbulas se extienden hacia adelante, dando a las ballenas su cabeza característica y larga, y los dos huesos más anteriores de la mandíbula superior (el maxilar y el premaxilar) están “telescopiados” hacia atrás, a veces cubriendo por completo la parte superior del cráneo. La migración posterior de estos huesos es el proceso mediante el cual las aberturas nasales se desplazaron a la parte superior del cráneo, creando espiráculos y desplazando el cerebro y el aparato auditivo hacia la parte posterior del cráneo. Los odontocetos (ballenas con dientes) tienen un solo espiráculo, mientras que los misticetos (ballenas con barbas) tienen espiráculos dobles. En los odontocetos, hay una asimetría pronunciada en los huesos telescopiados y en el espiráculo que brinda un medio natural de clasificación. Aunque los dientes aparecen a menudo en los misticetos fetales, solo los odontocetos exhiben dientes como adultos. Esos dientes son siempre conos o pernos simples; no se diferencian por región o función como los dientes en otros mamíferos. (Las ballenas no pueden masticar su alimento; en cambio, se tritura en un retículo, o “glandular crop”, muscular, que contiene piedras.) A diferencia del resto de los mamíferos, las ballenas no tienen glándulas lagrimales, no tienen glándulas cutáneas y no tienen sentido olfativo. Su audición es aguda pero el oído no tiene apertura externa. La audición ocurre a través de vibraciones transmitidas a un hueso grande y en forma de concha formado por la fusión de huesos del cráneo (el oído periótico y los bullas auditivos). Estos, entonces, son los rasgos principales de las ballenas. Algunos muestran claramente las adaptaciones distintivas impuestas a las ballenas por su compromiso con la vida marina; otros vinculan claramente a las ballenas con sus ancestros terrestres. Otros muestran las huellas de descenso desde un ancestro terrestre en común con varias especies antiguas y modernas. De todas estas características en conjunto, podemos reconstruir la vía que la evolución de las ballenas siguió desde un ancestro terrestre hasta una ballena moderna confinada a los océanos profundos. Pensando en la ascendencia de la ballena En 1693, John Ray registró su conclusión de que las ballenas son mamíferos basada en su similitud con mamíferos terrestres (Barnes 1984). La discusión científica pre-darwiniana giró en torno a si las ballenas descendían de mamíferos terrestres o eran ancestrales a ellos. Darwin (1859) sugirió que las ballenas surgieron de osos, esbozando un escenario en el que presiones selectivas podrían causar que los osos evolucionaran en ballenas; avergonzado por las críticas, retiró a los hipotéticos osos nadadores de ediciones posteriores del Origen (Gould 1995). Más tarde, Flower (1883) reconoció que las ballenas tienen rasgos rudimentarios persistentes y vestigiales característicos de los mamíferos terrestres, confirmando así que la dirección del descenso era de especies terrestres a especies marinas. Basándose en la morfología, Flower también vinculó las ballenas con los ungulados; parece haber sido la primera persona en hacerlo. A comienzos del siglo XX, Eberhard Fraas y Charles Andrews sugirieron que los creodontes (carnívoros primitivos, hoy extintos) eran los ancestros de las ballenas (Barnes 1984). Más tarde, WD Matthew del American Museum of Natural History postuló que las ballenas descendían de insectívoros, pero su idea nunca obtuvo mucho apoyo (Barnes 1984). Posteriormente, Everhard Johannes Slijper intentó combinar ambas ideas, afirmando que las ballenas descendían de lo que Barnes llamó acertadamente “creodontos-cum-insectívoros”. Sin embargo, nunca se ha encontrado un animal así. Más recientemente, Van Valen (1966) y Szalay (1969) asociaron las primeras ballenas con condilartos mesoníquidos (un grupo ya extinto de ungulados carnívoros primitivos, ninguno más grande que un lobo) sobre la base de caracteres dentarios. Evidencias más recientes confirman su evaluación. Así que Flower estuvo básicamente en lo cierto. La evidencia La evidencia de que las ballenas descendieron de mamíferos terrestres se divide aquí en nueve partes independientes: paleontológica, morfológica, biológica molecular, vestigial, embrionaria, geoquímica, paleoambiental, paleobiogeográfica y cronológica. Aunque mi resumen de la evidencia no es exhaustivo, muestra que la visión actual de la evolución de las ballenas está respaldada por investigación científica en varias disciplinas distintas. 1. Evidencia paleontológica La evidencia paleontológica proviene del estudio de la secuencia fósil desde mamíferos terrestres a formas cada vez más parecidas a ballenas hasta la aparición de las ballenas modernas. Aunque las primeras ballenas (arqueocetos) muestran mayor diversidad de la que tengo espacio para discutir aquí, los ejemplos en esta sección representan las tendencias que vemos en este taxón. Aunque hay dos subórdenes modernos de ballenas (Odontocetos y Misticetos), esta discusión se centrará en el origen de las ballenas como un orden de mamíferos y dejará de lado los problemas relacionados con la diversificación en subórdenes. Sinonyx Comenzamos con Sinonyx, un mesoníquido del tamaño de un lobo (un ungulado primitivo del orden Condylarthra, del que surgieron los artiodáctilos, perisodáctilos, proboscidios, etc.) del final del Paleoceno, hace unos 60 millones de años. Los caracteres que vinculan Sinonyx con las ballenas, indicando así que son parientes, incluyen un hocico alargado, un foramen yugular agrandado y un basicráneo corto (Zhou y otros 1995). El número de dientes era el número mamífero primitivo (44); los dientes estaban diferenciados como los heterodontes de los mamíferos actuales. Los molares eran dientes de corte muy estrechos, especialmente en la mandíbula inferior, pero poseían múltiples cúspides. La elongación del hocico suele asociarse a la caza de peces: todas las ballenas piscívoras, así como los delfines, tienen hocicos alargados. Estas características eran atípicas en los mesoníquidos, indicando que Sinonyx ya estaba desarrollando las adaptaciones que luego se volvieron la base de la forma de vida especializada de las ballenas. Pakicetus El siguiente fósil de la secuencia, Pakicetus, es el cetáceo más antiguo y el primer arqueoceto conocido. Es del eoceno temprano de Pakistán, de hace unos 52 millones de años (Gingerich y otros 1983). Aunque solo se conoce por restos craneales fragmentarios, esos restos son muy diagnósticos y son sin duda intermedios entre Sinonyx y ballenas posteriores. Esto es especialmente cierto para los dientes. Los molares superior e inferior, que tienen múltiples cúspides, siguen siendo similares a los de Sinonyx, pero los premolares se han convertido en dientes triangulares simples compuestos por una sola cúspide aserrada en sus bordes anterior y posterior. Los dientes de ballenas posteriores muestran aún más simplificación hacia triángulos simples aserrados, como los de tiburones carnívoros, indicando que los dientes de Pakicetus estaban adaptados para cazar peces.  Gingrich y otros (1983) publicaron esta reconstrucción del cráneo de Pakicetus inachus (redibujada para RNCSE por Janet Dreyer). Un cráneo bien conservado muestra que Pakicetus era sin duda un cetáceo con una caja craneal estrecha, una cresta sagital alta y estrecha y crestas lambdoideas prominentes. Gingerich y otros (1983) reconstruyeron un cráneo compuesto de unos 35 centímetros. Pakicetus no oía bien bajo el agua. Su cráneo no tenía ni bullas timpánicas densas ni senos que aislaran el área auditiva izquierda de la derecha, una adaptación de las ballenas posteriores que permite la audición direccional bajo el agua y evita la transmisión de sonidos a través del cráneo (Gingerich y otros 1983). Todas las ballenas actuales tienen senos llenos de espuma junto con bullas timpánicas densas que crean un contraste de impedancia para separar los sonidos que llegan desde direcciones distintas. En Pakicetus tampoco hay evidencia de vascularización del oído medio, necesaria para regular la presión en el oído medio durante la inmersión (Gingerich y otros 1983). Por lo tanto, Pakicetus probablemente era incapaz de realizar inmersiones de cualquier profundidad significativa. Esta evaluación paleontológica de la niche ecológica de Pakicetus es totalmente coherente con la evidencia geoquímica y paleoambiental. En cuanto a la audición, Pakicetus era más terrestre que acuático, pero la forma de su cráneo era definitivamente cetácea, y sus dientes se encontraban entre los estados ancestral y moderno.  Zhou y otros (1995) publicaron esta reconstrucción del cráneo de Sinonyx jiashanensis (redibujada para RNCSE por Janet Dreyer). Ambulocetus En el mismo lugar donde se encontró Pakicetus, pero en sedimentos unos 120 metros más altos, Thewissen y colegas (1994) descubrieron Ambulocetus natans, “la ballena caminante que nada”, en 1992. Datado en el Eoceno temprano a medio, de hace aproximadamente 50 millones de años, Ambulocetus es un fósil verdaderamente asombroso. Era claramente un cetáceo, pero también tenía patas funcionales y un esqueleto que aún permitía cierto grado de marcha terrestre. La conclusión de que Ambulocetus podía caminar usando las extremidades posteriores se apoya en su fémur grande y robusto. Sin embargo, como el fémur no tenía los puntos de inserción suficientemente grandes para los músculos de la marcha, no pudo haber sido un caminante muy eficiente. Probablemente solo podía caminar como lo hacen las focas modernas: rotando los pies posteriores hacia adelante y desplazándose por el suelo con ayuda de las extremidades anteriores y la flexión espinal. Al caminar, sus enormes patas delanteras debían apuntar lateralmente en buena medida, ya que, si apuntaban hacia adelante, se habrían interferido entre sí. Las extremidades anteriores también eran intermedias en estructura y función. El cúbito y el radio eran fuertes y capaces de soportar el peso del animal en tierra. El codo fuerte estaba inclinado hacia atrás, lo que permitía empujes posteriores del antebrazo para la natación. Sin embargo, las muñecas, a diferencia de las de las ballenas modernas, eran flexibles. Es obvio por la anatomía de la columna vertebral que Ambulocetus debió haber nadado con la espina vertebral oscilando arriba y abajo, propulsado por sus patas traseras orientadas hacia atrás. Como en otros mamíferos acuáticos que usan este método de natación, las patas traseras eran bastante grandes. De manera inusual, los dedos de las patas traseras terminaban en pezuñas, dando a conocer la ascendencia de ungulados del animal. La única vértebra caudal hallada es larga, lo que sugiere que la cola también era larga. Las vértebras cervicales eran relativamente largas en comparación con las de las ballenas modernas; Ambulocetus debió tener un cuello flexible. El cráneo de Ambulocetus era bastante cetáceo (Novacek 1994). Tenía un hocico largo, dientes muy similares a los de arqueocetos posteriores, un arco cigomático reducido y un bulla timpánica (que sostiene el tímpano) que estaba mal adherida al cráneo. Aunque Ambulocetus aparentemente carecía de espiráculo, las demás características craneales califican a Ambulocetus como cetáceo. Las características poscraniales están claramente en adaptación transicional al medio acuático. Así, Ambulocetus se describe mejor como un anfibio de tamaño similar a un león marino y comedor de peces que aún no estaba totalmente desconectado de la vida terrestre de sus ancestros. Rodhocetus En el Eoceno medio (46-47 millones de años) Rodhocetus llevó todos estos cambios aún más lejos, y aun así retuvo varias características terrestres primitivas (Gingerich y otros 1994). Es el arqueoceto más antiguo del cual se han conservado todas las vértebras torácicas, lumbares y sacras. Las vértebras lumbares tenían espinas neurales más altas que en ballenas anteriores. El tamaño de estas extensiones en la parte superior de las vértebras donde se insertan músculos indica que Rodhocetus había desarrollado una cola potente para nadar. 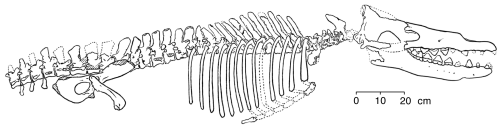 Gingrich y otros (1994) publicaron esta reconstrucción del esqueleto de Rodhocetus kasrani (redibujada para RNCSE por Janet Dreyer). En otra parte de la columna vertebral, las cuatro grandes vértebras sacras no estaban fusionadas. Esto daba más flexibilidad a la columna y permitía un empuje más potente al nadar. También es probable que Rodhocetus tuviera una aleta de cola, aunque tal rasgo no se conserva en los fósiles conocidos: poseía rasgos —cervicales acortadas, vértebras caudales proximales pesadas y robustas, y grandes espinas dorsales en las vértebras lumbares para grandes inserciones de músculos caudales y otros músculos axiales— que en ballenas modernas se asocian con el desarrollo y uso de aletas de cola. En conjunto, Rodhocetus debió haber sido un excelente nadador de cola, y es la ballena fósil más antigua comprometida con esta forma de natación. La pelvis de Rodhocetus era más pequeña que la de sus predecesores, pero aún estaba conectada a las vértebras sacras, lo que significa que Rodhocetus todavía podía caminar en tierra en cierta medida. Sin embargo, el ilion de la pelvis era corto en comparación con el de los mesoníquidos, lo que producía un empuje muscular menos potente desde la cadera durante la marcha, y el fémur era aproximadamente 1/3 más corto que el de Ambulocetus; por ello Rodhocetus probablemente no se movía tan bien en tierra como sus predecesores (Gingerich y otros 1994). El cráneo de Rodhocetus era bastante grande en comparación con el resto del esqueleto. Los premaxilares y la mandíbula estaban extendidos hacia adelante aún más que en sus predecesores, alargando el cráneo y haciéndolo todavía más cetáceo. Los molares tienen coronas más altas que en ballenas anteriores y están muy simplificados. Los molares inferiores son más altos que anchos. Hay una diferenciación reducida entre los dientes. Por primera vez, las fosas nasales se desplazaron hacia atrás a lo largo del hocico y se ubican encima de los dientes caninos, mostrando la evolución del espiráculo. Las bullas auditivas son grandes y de hueso denso (características únicas de cetáceos), pero aparentemente no contenían los senos típicos de ballenas posteriores, lo que hace cuestionable si Rodhocetus poseía audición direccional bajo el agua. En general, Rodhocetus mostró mejoras sobre ballenas anteriores por su tórax profundo y delgado, cabeza más larga, mayor flexibilidad vertebral y musculatura caudal expandida. El aumento de flexibilidad y fuerza en la espalda y cola con la disminución simultánea de fuerza y tamaño de las extremidades indicó que era un buen nadador de cola con menor capacidad para caminar en tierra. Basilosaurus La bien conocida ballena fósil Basilosaurus representa el siguiente grado evolutivo en la evolución de las ballenas (Gingerich 1994). Vivió durante el Eoceno tardío y la parte más reciente del Eoceno medio (35-45 millones de años atrás). Basilosaurus era un animal largo, delgado y serpentiforme que inicialmente se pensó que eran restos de una serpiente marina (de ahí su nombre, que realmente significa “lagarto rey”). Su longitud corporal extrema (de unos 15 metros) parece deberse a una característica única entre las ballenas; sus 67 vértebras son tan largas en comparación con otras ballenas de la época y con las ballenas modernas que probablemente representa una especialización que lo distingue de la línea que dio origen a las ballenas modernas. Lo que hace a Basilosaurus una ballena especialmente interesante, sin embargo, es la anatomía distintiva de sus extremidades posteriores (Gingerich y otros 1990). Tenía una cintura pélvica casi completa y un conjunto de huesos de las extremidades posteriores. Las extremidades eran demasiado pequeñas para una propulsión efectiva, de menos de 60 cm de longitud en este animal de 15 metros, y la pelvis estaba completamente aislada de la columna vertebral de modo que era imposible soportar peso. Las reconstrucciones del animal han situado sus patas externas al cuerpo, una configuración que representaría una forma intermedia importante en la evolución de las ballenas. Aunque nunca se ha encontrado ninguna aleta de cola (puesto que las aletas de cola no contienen huesos y es poco probable su fosilización), Gingerich y otros (1990) señalaron que la columna vertebral de Basilosaurus comparte características de ballenas que sí tienen aletas de cola. La cola y las vértebras cervicales son más cortas que las de las regiones torácica y lumbar, y Gingerich y otros (1990) toman estas proporciones vertebrales como evidencia de que Basilosaurus probablemente también tenía una aleta de cola. Otra evidencia de que Basilosaurus pasó la mayor parte de su tiempo en el agua proviene de otro cambio importante en el cráneo. Este animal tenía una gran abertura nasal única que había migrado una corta distancia hacia atrás hasta una posición correspondiente al tercio posterior del conjunto dental. El movimiento desde el extremo anterior del hocico hacia una posición más cerca de la parte superior de la cabeza es característico solo de aquellos mamíferos que viven en ambientes marinos o acuáticos. Dorudon Dorudon fue contemporáneo de Basilosaurus en el Eoceno tardío (aproximadamente 40 millones de años atrás) y probablemente representa el grupo más probable como ancestral de las ballenas modernas (Gingerich 1994). Dorudon carecía de las vértebras alargadas de Basilosaurus y era mucho más pequeño (de unos 4-5 metros de longitud). La dentición de Dorudon era similar a la de Basilosaurus; su cráneo, comparado con los cráneos de Basilosaurus y las ballenas previas, era algo abovedado (Kellogg 1936). Dorudon tampoco tenía todavía la anatomía craneal que indica la presencia del aparato necesario para la ecolocalización (Barnes 1984).  Gingrich y Uhen (1996) publicaron esta reconstrucción del esqueleto de Dorudon atrox (redibujada para RNCSE por Janet Dreyer). Basilosaurus y Dorudon eran ballenas plenamente acuáticas (como Basilosaurus, Dorudon tenía extremidades posteriores muy pequeñas que quizás apenas sobresalían de la pared corporal). Ya no estaban ligadas a la tierra; de hecho, no habrían podido moverse en tierra en absoluto. Su tamaño y la ausencia de extremidades que soportaran su peso las convirtió en mamíferos obligatoriamente acuáticos, una tendencia que se desarrolla y refuerza en taxones de ballenas posteriores. Con claridad, incluso si solo miramos la evidencia paleontológica, la afirmación creacionista de “¡No hay intermediarios fósiles!” está equivocada. De hecho, en el caso de las ballenas tenemos varios, ordenados bellamente en orden morfológico y cronológico. Al resumir la evidencia paleontológica, hemos señalado los cambios consistentes que indican una serie de adaptaciones de entornos más terrestres a más acuáticos a medida que avanzamos de la especie más ancestral a la más reciente. Estos cambios afectan la forma del cráneo, la forma de los dientes, la posición de las fosas nasales, el tamaño y la estructura tanto de las extremidades anteriores como posteriores, el tamaño y forma de la cola y la estructura del oído medio en relación con la audición direccional bajo el agua y la inmersión. La evidencia paleontológica registra una historia de adaptación creciente a la vida en el agua, no a cualquier forma de vida en el agua, sino a la vida vivida por las ballenas contemporáneas. 2. Evidencia morfológica La revisión de las características morfológicas compartidas por ballenas fósiles y ungulados vivos hace aún más clara su descendencia común. Por ejemplo, la anatomía del pie de Basilosaurus vincula a las ballenas con los artiodáctilos (Gingerich y otros 1990). El eje de simetría del pie en estas ballenas fósiles cae entre el tercer y cuarto dedos. Esta disposición se llama paraxónica y es característica de artiodáctilos, ballenas y condílaros, y se halla raramente en otros grupos (Wyss 1990). Otro ejemplo involucra el yunque (el “yunque” del oído medio). El yunque de Pakicetus, conservado al menos en un ejemplar, es morfológicamente intermedio en todos sus caracteres entre el yunque de las ballenas modernas y el de los artiodáctilos modernos (Thewissen y Hussain 1993). Además, la articulación entre el martillo (malleus) y el yunque de la mayoría de mamíferos se orienta en un ángulo entre el medio y el frente del animal (rostromedialmente), mientras que en las ballenas modernas y en ungulados se orienta en un ángulo entre el lado y el frente (rostrolateralmente). En Pakicetus, el primer fósil cetáceo, la articulación está orientada rostralmente (intermedia en posición entre las condiciones ancestral y derivada). Así, la articulación claramente ha rotado hacia la región media desde la condición ancestral en mamíferos terrestres (Thewissen y Hussain 1993); Pakicetus nos proporciona una instantánea de la transición. 3. Evidencia biológica molecular La hipótesis de que las ballenas descienden de mamíferos terrestres predice que las ballenas vivas y mamíferos terrestres vivos estrechamente relacionados muestren similitudes en su biología molecular aproximadamente en proporción a la reciente descendencia común. Es decir, las ballenas deberían parecerse más en su biología molecular a grupos de animales con los que comparten un ancestro más reciente que a otros animales que exhiben similitudes convergentes en morfología, ecología o comportamiento. En contraste, el creacionismo no tiene base científica para predecir cuáles deben ser los patrones de similitud, porque no hay forma científica de predecir cómo el creador decidió distribuir similitudes moleculares entre especies. Estudios moleculares de Goodman y otros (1985) muestran que las ballenas están más estrechamente relacionadas con ungulados que con cualquier otro mamífero, un resultado coherente con las expectativas evolutivas. Estos estudios examinaron mioglobina, cristalina alpha del cristalino y citocromo c en un estudio de 46 especies diferentes de mamíferos. Miyamoto y Goodman (1986) ampliaron luego el número de secuencias proteicas al incluir hemoglobinas alfa y beta y ribonucleasa; también aumentaron el número de mamíferos incluidos al estudio a 72. Los resultados fueron los mismos: las ballenas están claramente incluidas entre los ungulados. Otros estudios moleculares de diversos genes, proteínas y enzimas por Irwin y otros (1991), Irwin y Arnason (1994), Milinkovitch (1992), Graur y Higgins (1994), Gatesy y otros (1996) y Shimamura y otros (1997) también identificaron a las ballenas como estrechamente relacionadas con artiodáctilos, aunque haya diferencias en los detalles entre los estudios. Al ubicar a las ballenas cerca e incluso firmemente dentro de los artiodáctilos, estos estudios moleculares confirman las predicciones hechas por la teoría evolutiva. Este patrón de similitudes bioquímicas debe estar presente si las ballenas y los ungulados, especialmente los artiodáctilos, comparten un ancestro común cercano. El hecho de que estas similitudes estén presentes es por lo tanto una evidencia sólida de la descendencia común de ballenas y ungulados. 4. Evidencia vestigial Las características vestigiales de las ballenas nos dicen dos cosas. Nos dicen que las ballenas, como tantos otros organismos, tienen rasgos que no tienen sentido desde una perspectiva de diseño: no tienen función actual, requieren energía para producirse y mantenerse, y pueden ser perjudiciales para el organismo. También nos dicen que las ballenas llevan consigo un fragmento de su pasado evolutivo, destacando una historia de ascendencia terrestre. Las ballenas modernas suelen conservar vestigios en forma de varillas de huesos pélvicos, fémures y tibias, todos incrustados en la musculatura de sus paredes corporales. Estos huesos son más pronunciados en especies anteriores y menos pronunciados en especies posteriores. Como muestra el ejemplo de Basilosaurus, las ballenas de edad intermedia tienen pelves vestigiales e extremidades posteriores de tamaño intermedio. Las ballenas también conservan un número de estructuras vestigiales en sus órganos sensoriales. Las ballenas modernas solo tienen nervios olfatorios vestigiales. Además, en las ballenas modernas el meato auditivo (la apertura exterior del conducto del oído) está cerrado. En muchas, es apenas del tamaño de un fino filamento, de unos 1 mm de diámetro, y a menudo se estrangula a mitad de camino. Todas las ballenas tienen varios músculos pequeños dedicados a oídos externos inexistentes, que aparentemente son vestigios de un tiempo en que podían mover sus orejas, un comportamiento que usan típicamente los animales terrestres para la audición direccional. El diafragma en las ballenas es vestigial y tiene muy poco músculo. Las ballenas usan el movimiento hacia fuera de las costillas para llenar sus pulmones de aire. Finalmente, Gould (1983) informó varias observaciones de cachalotes capturados con extremidades posteriores visibles y protuberantes. Del mismo modo, se han visto delfines con pequeñas aletas pélvicas, aunque probablemente no estaban soportadas por huesos de extremidades como en esos raros cachalotes. Y algunas ballenas, como las belugas, poseen pinnas auditivas rudimentarias, una característica que no puede servir a ningún propósito en un animal sin oído externo y que puede reducir la eficiencia de nado al aumentar la resistencia hidrodinámica durante la natación. Aunque esta lista no es ni por asomo exhaustiva, queda claro que las ballenas tienen una gran cantidad de rasgos vestigiales heredados de sus ancestros terrestres. 5. Evidencia embrionaria Como las características vestigiales, las características embrionarias también nos dicen dos cosas. Primero, el embrión de la ballena desarrolla una serie de rasgos que más tarde abandona antes de alcanzar su forma final. ¿Cómo puede explicar el creacionismo un proceso aparentemente sin sentido, edificando estructuras solo para abandonarlas o destruirlas más tarde? Darwin (1859) planteó la misma pregunta. ¿No tendría más sentido que los embriones alcanzaran sus formas adultas rápidamente y directamente? Parece poco razonable para un diseñador o creador perfecto enviar al embrión por un camino tan tortuoso, pero la evolución requiere que las nuevas características se construyan sobre la base de características anteriores que luego modificaría o descartaría. Segundo, la embriología de la ballena, examinada en detalle, también aporta evidencia de su ascendencia terrestre. Como embriones y no menos que como animales adultos, las ballenas son vertederos, por decirlo así, de rasgos viejos descartados que ya no les sirven. Muchas ballenas, mientras aún están en el útero, empiezan a desarrollar vello corporal. Sin embargo, ninguna ballena moderna conserva vello corporal tras el nacimiento, excepto algunas vibrisas del hocico y pelos alrededor de sus espiráculos usados como cerdas sensoriales en pocas especies. El hecho de que las ballenas posean los genes para producir vello corporal muestra que sus ancestros tenían vello corporal. En otras palabras, sus ancestros eran mamíferos corrientes. En muchos embriones de ballenas, los brotes de extremidad posterior son visibles por un tiempo, pero desaparecen a medida que la ballena crece. También son visibles en el embrión las orejas pinna rudimentarias, que desaparecen antes del nacimiento (excepto en aquellos que las portan como atavismos raros). Y, en algunas ballenas, los lóbulos olfatorios del cerebro existen solo en el feto. El embrión de la ballena comienza con las fosas nasales en el lugar habitual de los mamíferos, en la punta del hocico. Pero durante el desarrollo, las fosas nasales migran a su lugar final en la parte superior de la cabeza para formar el espiráculo (o espiráculos). También podemos comprender la evolución dentro de las ballenas mediante su embriología. Sabemos que los misticetos evolucionaron a partir de los odontocetos: algunos embriones de misticetos comienzan a desarrollar dientes. Como con el vello corporal, los dientes desaparecen antes del nacimiento. Dado que no hay utilidad de dientes en el útero, solo la herencia de un ancestro común tiene sentido; no hay razón para que el diseñador inteligente o creador especial provea dientes a embriones de ballena. Así tenemos aún otro campo independiente en completo acuerdo con la tesis general: las ballenas poseen rasgos que las conectan con ancestros mamíferos terrestres, en particular los mamíferos con pezuñas. 6. Evidencia geoquímica Las primeras ballenas vivieron en hábitats de agua dulce, pero los ancestros de las ballenas modernas se trasladaron a hábitats de agua salada y por ello tuvieron que adaptarse a beber agua salada. Como agua dulce y agua salada tienen relaciones isotópicas de oxígeno algo diferentes, podemos predecir que la transición quedará registrada en los restos esqueléticos de las ballenas, cuyos más duraderos son los dientes. Como era de esperar, los dientes fósiles de las ballenas más antiguas tienen relaciones de oxígeno pesado a oxígeno ligero más bajas, indicando que los animales bebían agua dulce (Thewissen y otros 1996). Los dientes fósiles de ballenas posteriores tienen relaciones de oxígeno pesado a oxígeno ligero más altas, indicando que bebían agua salada. Esto refuerza absolutamente la inferencia derivada de toda la otra evidencia discutida aquí: los ancestros de las ballenas modernas se adaptaron de hábitats terrestres a hábitats de agua salada a través de hábitats de agua dulce. 7. Evidencia paleoambiental La evolución hace otras predicciones sobre la historia de los taxones basadas en la visión de conjunto de los fósiles en un contexto ambiental más amplio. La secuencia de fósiles de ballenas y sus cambios también debería relacionarse con cambios observados en los registros fósiles de otros organismos al mismo tiempo y en entornos similares. Los fósiles de otros organismos asociados con los fósiles de ballenas indican el ambiente en el que vivían las ballenas. Además, esta evidencia debería ser coherente con la evidencia de otras áreas de estudio. Deberíamos esperar encontrar evidencia de una serie de entornos transicionales, de lo plenamente terrestre a lo plenamente marino, ocupados por la serie de especies de ballenas del registro fósil. La morfología de Sinonyx indica que era completamente terrestre. No debe sorprender, por tanto, que sus fósiles se encuentren asociados con los fósiles de otros animales terrestres. Pakicetus probablemente pasó gran parte del tiempo en el agua en busca de alimento. Aunque la fauna mamífera encontrada con Pakicetus consiste en roedores, murciélagos, varios artiodáctilos, perisodáctilos y proboscídeos e incluso un primate (Gingerich y otros 1983), también hay animales acuáticos como caracoles, peces, tortugas y cocodrilos. Además, el sedimento asociado con Pakicetus muestra evidencia de corriente o flujo, usualmente asociada con suelos transportados por agua. La evidencia paleoambiental muestra así claramente que Pakicetus vivía en un entorno terrestre bajo en elevación y húmedo, con excursiones ocasionales al agua dulce. Curiosamente, tanto los dientes deciduos como los permanentes del animal se encuentran en esos sedimentos con una frecuencia similar, lo que apoya la idea de que Pakicetus daba a luz en tierra. Los sedimentos donde se encontró Ambulocetus contienen impresiones de hojas además de fósiles del caracol espiral Turritella y otros moluscos marinos. Claramente, la presencia de tales fósiles debe significar que el fósil de Ambulocetus se encontró en lo que alguna vez fue un mar somero, aunque las hojas pueden arrastrarse al mar y fosilizarse allí, los moluscos marinos no se encontrarían en tierra. Rodhocetus se encuentra en pizarras verdes depositadas en la zona nerítica profunda (equivalente a la parte externa del talud continental). Debido a que las pizarras verdes se asocian con aguas de fondo de baja oxigenación, Rodhocetus debió vivir a mayor profundidad que cualquier cetáceo anterior. El hecho de que se encuentre asociado con foraminíferos planctónicos y otros microfósiles concuerda con esta determinación de la profundidad del agua. Basilosaurus y Dorudon han sido hallados en una variedad de tipos de sedimento (Kellogg 1936), lo que indica que eran de amplia distribución y capaces de vivir en aguas profundas así como someras. A partir de la evidencia paleoambiental, podemos ver claramente que, a medida que las ballenas evolucionaban, avanzaban hacia aguas más profundas y se liberaban progresivamente de los ambientes terrestres y costeros. 8. Evidencia paleobiogeográfica La evidencia geográfica también es coherente con los patrones de distribución esperados para la primera aparición de las ballenas y su expansión geográfica posterior. Esperaríamos que las especies terrestres tuvieran una distribución geográfica más restringida que las especies marinas, que tienen esencialmente el océano entero como rango geográfico. El rango de Sinonyx está restringido al Asia central. Los ejemplares de Pakicetus solo se han encontrado en Pakistán; Ambulocetus y Rodhocetus parecen tener una restricción similar. En contraste, Basilosaurus y Dorudon, que representan a las ballenas más adaptadas a la vida en mar abierto, se hallan en una zona mucho más amplia. Sus fósiles se han encontrado tan lejos de Asia meridional como Georgia, Luisiana y la Columbia Británica. Durante el Eoceno, la mayoría de las áreas en las que se han encontrado fósiles de las ballenas posteriores estaban bastante cercanas entre sí. De hecho, la mayoría se sitúa a lo largo del borde externo de un mar antiguo llamado Tethys, cuyos restos hoy son el Mediterráneo, el Caspio, el Negro y el Aral. La distribución biogeográfica de ballenas fósiles coincide con el patrón predicho por la evolución: las ballenas se encuentran inicialmente en un área geográfica bastante pequeña y no se distribuyeron por todo el mundo hasta después de haber evolucionado en animales plenamente acuáticos que ya no estaban atados a la tierra. 9. Evidencia cronológica El último tramo de evidencia en nuestra imagen mutuamente coherente sobre los orígenes de las ballenas proviene de una consideración de por qué las ballenas se originaron cuando lo hicieron. La evolución es una respuesta a retos y oportunidades ambientales. Durante el Cenozoico temprano, a los mamíferos se les ofreció un nuevo conjunto de oportunidades de radiación y diversificación debido, en parte, al vacío dejado por extinciones masivas al final del Período Cretácico. Como los reptiles ya no predominaban, surgieron nuevas formas en que los mamíferos podían sobrevivir. En el caso específico de las ballenas, los reptiles nadadores de los océanos del mundo ya no podían mantener a los mamíferos a distancia. Antes de las extinciones del Cretácico tardío, los reptiles marinos mesozoicos como los plesiosaurios, ictiosaurios, mosasaurios y cocodrilos marinos probablemente habrían disfrutado de cualquier mamífero que se apartara de la costa en busca de alimento. Una vez que esos depredadores desaparecieron, la evolución produjo rápidamente mamíferos, incluidas las ballenas, que estaban tan a gusto en los mares como antes en la tierra. La transición tomó unos 10-15 millones de años para producir ballenas plenamente acuáticas y de buceo profundo con audición direccional bajo el agua. La evolución predice que las ballenas no pudieron aparecer y radiarse con éxito antes del Eoceno, y que los mamíferos debieron radiarse a ambientes marinos como lo hicieron hacia una gran variedad de otros ambientes vaciados por los reptiles al final del Cretácico. Conclusión Tomado en conjunto, toda esta evidencia apunta a una sola conclusión: que las ballenas evolucionaron a partir de mamíferos terrestres. Hemos visto que existen nueve áreas de estudio independientes que aportan evidencia de que las ballenas comparten un ancestro común con mamíferos con pezuñas. La fuerza de la evidencia de áreas de estudio independientes que apoyan la misma conclusión hace que sea totalmente irrazonable la refutación por escenarios de creación especial, incredulidad personal, el argumento de ignorancia o escenarios de “diseño inteligente”. La única conclusión científica plausible es que las ballenas sí evolucionaron de mamíferos terrestres. Así que no importa cuánto se quejen los antievolucionistas sobre lo imposible que sería que mamíferos terrestres vellosos se conviertan en ballenas completamente acuáticas, la evidencia misma los desmiente. Este es el poder de usar líneas de evidencia mutuamente reforzadas e independientes. Espero que se convierta en un arma principal para derribar objeciones antievolucionistas infundadas y apoyar el pensamiento evolutivo en el público en general. Así funciona la ciencia real, y debemos enfatizar el proceso de inferencia científica a medida que señalamos las conclusiones que los científicos extraen de la evidencia: que las predicciones concordantes de campos independientes de estudio científico confirman el mismo patrón de ascendencia de las ballenas. Agradecimientos Me gustaría agradecer al Dr. Philip Gingerich por su ayuda y revisión de este artículo. Referencias Barnes LG. Search for the first whale. Oceans 1984 March-April; 17 (2): 20-3. Darwin CR. On the Origin of Species. New York: Random House, 1859. Flower WH. On the arrangement of the orders and families of existing Mammalia. Proceedings of the Zoological Society of London 1883 Aug; 1: 178-86. Gatesy J, Hayashi C, Cronin MA, Arctander P. Evidence from milk casein genes that cetaceans are close relatives of hippopotamid artiodactyls. Molecular Biology and Evolution 1996; 13 (7): 954-63. Gingerich P. The whales of Tethys. Natural History 1994 April; 103 (4): 86-8. Gingerich P, Raza SM, Arif M, Anwar M, Zhou X. New whale from the Eocene of Pakistan and the origin of cetacean swimming. Nature 1994; 368: 844-7. Gingerich P, Smith BH, Simons EL. Hind limbs of Eocene Basilosaurus: evidence of feet in whales. Science 1990; 249: 154-7. Gingerich P, Wells NA, Russell DE, Shah SMI. Origin of whales in epicontinental remnant seas: new evidence from the early Eocene of Pakistan, Science 1983; 220: 403-6. Gish DT. Evolution: The Challenge of the Fossil Record. El Cajon (CA): Creation-Life Publishers, 1985. Goodman M, Czelusniak J, Beeber J. Phylogeny of primates and other eutherian orders: a cladistic analysis using amino acid and nucleotide sequence data. Cladistics 1985; 1 (2): 171-85. Gould SJ. Hen's teeth and horse's toes. In: Hen's Teeth and Horse's Toes. Norton: New York, 1983. p 177-86. Gould SJ. Hooking leviathan by its past. In: Dinosaur in a Haystack, New York: Harmony Books, 1995. p 359-76. Graur D, Higgins DG. Molecular evidence for the inclusion of cetaceans within the order Artiodactyla. Molecular Biology and Evolution 1994; 11 (3): 357-64. Irwin DM, Arnason U. Cytochrome b gene of marine mammals: phylogeny and evolution. Journal of Mammalian Evolution 1994; 2 (1): 37-55. Irwin DM, Kochner TD, Wilson AC. Evolution of the cytochrome b gene of mammals. Journal of Molecular Evolution 1991; 32: 128-44. Kellogg R. A Review of the Archaeoceti. Washington DC: Carnegie Institute, 1936. Matthews LH. The Natural History of the Whale. New York: Columbia University Press, 1978. Milinkovitch MC. DNA-DNA hybridizations support ungulate ancestry of cetacea. Journal of Evolutionary Biology 1992; 5: 149-60 Milinkovitch MC, Orti G, Meyer A. Revised phylogeny of whales suggested by mitochondrial ribosomal DNA sequences. Nature 1993; 361: 346-8. Miyamoto MM, Goodman M. Biomolecular systematics of eutherian mammals: phylogenetic patterns and classification. Systematic Zoology 1986; 35 (2):230-40. Novacek M. Mammalian phylogeny: shaking the tree. Nature 1992; 356: 121-5. Novacek M. Whales leave the beach. Nature 1994; 368: 807. Shimamura M, Yasue H, Ohshima K, Abe H, Kato H, Kishiro T, Goto M, Munechika I, Okada N. Molecular evidence from retroposons that whales form a clade within even-toed ungulates. Nature 1997; 388: 666-9. Szalay FS. Origin and evolution of function of the mesonychid condylarth feeding mechanism. Evolution 1969; 23: 703-20. Thewissen JGM, Hussain ST. Origin of underwater hearing in whales. Nature 1993; 361: 444-5. Thewissen JGM, Hussain ST, Arif M. Fossil evidence for the origin of aquatic locomotion in Archaeocete whales. Science 1994; 263: 210-2. Thewissen JGM, Roe LJ, O'Neill JR, Hussain ST, Sahni A, Bajpal S. Evolution of cetacean osmoregulation. Nature 1996; 381: 379-80. Van Valen L. Deltatheridia, a new order of mammals. Bulletin of the American Museum of Natural History 1966; 132: 1-126. Wyss A. Clues to the origin of whales. Nature 1990; 347: 428-9. Zhou X, Zhai R, Gingerich P, Chen L. Skull of a new Mesonychid (Mammalia, Mesonychia) from the late Paleocene of China. Journal of Vertebrate Paleontology 1995; 15 (2): 387-400. Dirección del autor: Ray Sutera 81 Franklin Ave. Ocean Grove NJ 07756-1128 rsutera@aol.com |