Tudo sobre Archaeopteryx
por Chris Nedin[Referências Editadas: 10 de junho de 2002]
![]()
Sumário
- Introdução
- Espécimes de Archaeopteryx
- Características de Archaeopteryx
- Archaeopteryx + aves modernas, apenas variação dentro da espécie?
- Ratitas
- A história de dois ossos pélvicos
- Volar ou não voar
- A ancestralidade de Archaeopteryx
- Protoavis
- Conclusões
- Agradecimentos
- Referências
Introdução
Archaeopteryx lithographica ("asa antiga da pedra de impressão").
Nomeado após o calcário no qual foi descoberto. A pedra é um calcário liso e de grão fino que foi utilizado na impressão. Extraído de e ao redor da área de Solnhofen na Alemanha. Formado no fundo de uma lagoa hipersalina no Jurássico Superior, há cerca de 150 milhões de anos.
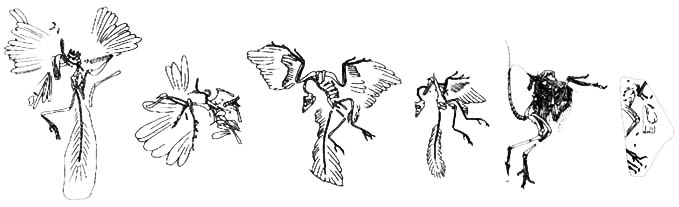
Archaeopteryx espécimes
Foram encontrados 8 espécimes de Archaeopteryx (7 espécimes reais e uma pena). Essas descobertas estão documentadas cronologicamente (por descrição) abaixo.
- A Peninha
Encontrada em 1860 perto de Solnhofen e foi uma revelação quando foi descrita por H. v Meyer em 1861. A surpresa não foi a idade do fóssil, já que várias pegadas de dinossauros ornitópodes erroneamente atribuídas a aves eram conhecidas do Triássico, mas o detalhe que foi preservado.
- O Especime de Londres

Encontrado em 1861, perto de Langenaltheim. Provavelmente o mais conhecido (juntamente com o espécime de Berlim). Sua descoberta foi anunciada por H. v Meyer em 1861 e o espécime foi posteriormente comprado pelo Museu Britânico de História Natural em Londres (sob a instrução de Richard Owen). Custou 700 libras esterlinas - uma pequena fortuna naqueles tempos, mas por aquele preço Owen também recebeu pouco mais de mil outros fósseis de Solnhofen. O espécime foi vendido pelo colecionador amador e médico local Carl Haberlein, que o havia recebido em lugar de pagamento por tratamento médico. Owen descreveu o espécime em 1863. Ele viu imediatamente que era uma descoberta importante e reconheceu que representava uma forma transicional - mas não no sentido de "Darwin". Owen era um firme "evolucionista", no entanto ele não acreditava no modelo de evolução de Darwin. Curiosamente, Huxley, que era um firme "darwinista", falhou em reconhecer a verdadeira importância do fóssil e meramente comentou sobre ele como um "ave semelhante a réptil". Não foi até comparações próximas serem feitas com o dinossauro Compsognathus que o verdadeiro valor de Archae foi realizado. - O Especime de Berlim

Encontrado em 1877 perto de Blumenberg. Este era um espécime melhor que o de Londres, principalmente porque tinha uma cabeça completa, embora muito amassada, e foi rapidamente adquirido pelo museu de Berlim. Foi vendido a eles pelo filho de Carl Haberlein (falar sobre mantê-lo na família!). Foi descrito por W. Dames em 1884.
- O Especime de Maxburg

Encontrado em 1958 perto de Langenaltheim (o mesmo que o Especime de Londres). Este espécime é apenas do tronco e é o único espécime que ainda está em mãos privadas. Em 1992, após a morte de seu descobridor e proprietário Eduard Opitsch, o espécime foi encontrado desaparecido e acredita-se que tenha sido vendido secretamente (Abbott 1992). Seu paradeiro permanece desconhecido. O espécime foi descrito por Heller em 1959.
- O Especime de Haarlem ou Teyler

Este espécime foi na verdade encontrado perto de Reidenburg em 1855, 5 anos antes da peninha! Ele estava em um museu após ser classificado como Pterodactylus crassipes por H. v Meyer em 1875. Curiosamente, Mayer o descreveu como tendo uma membrana de voo diferente de qualquer outro pterodáctilo conhecido, agora sabemos o porquê! Uma reexaminação do fóssil em 1970 por Ostrom revelou penas e sua verdadeira identidade.
- O Especime de Eichstatt

Encontrado perto de Workerszell em 1951, foi descrito por P. Wellnhofer em 1974. Este é o menor de todos os espécimes, sendo cerca de 2/3 do tamanho dos outros. Ele também difere em outros aspectos, como a estrutura dos dentes e os ossos do ombro mal ossificados. Sugeriu-se que este é um gênero separado, no entanto as diferenças também podem ser atribuídas à possível fase juvenil do animal e/ou a um nicho alimentar diferente. No entanto, este espécime tem a cabeça melhor preservada, da qual a litania de características cranianas reptilianas de Archae foram descritas. No momento, ele ainda reside dentro de A. lithographica.
- O Especime de Solnhofen

Encontrado na década de 1960 perto de Eichstatt por um trabalhador turco. Primeiro identificado como Compsognathus, por um colecionador amador, no entanto, exames posteriores mostraram que os braços eram muito longos para o tamanho do corpo e a preparação revelou traços de penas. Descrito por P. Wellnhofer em 1988.
- O Especime Solnhofen-Aktien-Verein
Um novo espécime foi descrito por Wellnhofer (1993), mas a descrição está em alemão e, portanto, as informações são limitadas. O espécime tem sido classificado como uma nova espécie, Archaeopteryx bavarica, e tem sido relatado como possuindo um esterno ossificado pequeno, bem como impressões de penas.
Características do Archaeopteryx
Muito tem sido dito nos círculos pseudocientíficos sobre a posição do Archae no esquema evolutivo das coisas. O "argumento" usual apresentado é que o Archae não pode ser um fóssil transicional entre aves e dinossauros porque é uma ave. Essa linha simplista contradiz o fato de que, embora o Archae seja de fato classificado como uma ave, essa classificação foi feita com base em 4 principais características - 2 das quais não são exclusivas das aves. Essa classificação ignora o fato de que o Archae possui numerosas características que são exclusivas, exclusivas no sentido de que elas não são possuídas por aves. As afinidades avianas do Archae são aceitáveis com base nas seguintes 4 principais características:
Características avianas do Archaeopteryx
- 1) Penas.
-
As penas são a característica diagnóstica dos pássaros modernos. Este é um dos principais critérios para classificar o Archae como um pássaro, pois nenhum outro animal moderno possui penas. A posse de penas é uma característica dos pássaros, então marque um ponto a favor dos pássaros. No entanto, no final de 1996, uma descoberta na China pode mudar essa visão. Um pequeno dinossauro terópode Sinosauropteryx (Chen et al. 1998) foi encontrado com o que parece ser penas preservadas ao longo da costas. A identificação das estruturas é ambígua, no entanto, (por exemplo, Unwin 1998), com alguns duvidando que as estruturas sejam penas.
PARA LER AGORA
Dinossauros com Penas Encontrados
Dois espécies de dinossauros foram recentemente encontradas no nordeste da China que possuem penas (Qiang et al. 1998). Protoarchaeopteryx robusta e Caudipteryx zoui mostram régeges, rectrizes e impressões de penas plumosas. Além disso, eles não são pássaros, carecendo de um dedão grande revertido (voltado para trás) (veja o número 2 abaixo) e um contato quadrijugal escamosal, tendo um quadrijugal unido ao quatrate por um ligamento e um processo do ísquio reduzido ou ausente. Estes e outros caracteres agrupam Protoarchaeopteryx e Caudipteryx com coelurosaurianos maniraptóides em vez de pássaros.
[Nota de Sistemática (de Padian 1998): Os sistemáticos definem os nomes de organismos por sua ancestralidade, neste caso os pássaros (Aves) consistem em Archaeopteryx mais pássaros vivos e todos os descendentes de seu ancestral comum mais recente. Os pássaros são diagnosticados por características únicas que apenas eles possuem e que são herdadas desse ancestral comum. Mesmo se as penas forem compartilhadas por um grupo mais amplo do que apenas pássaros, os pássaros ainda são definidos como Archaeopteryx e parentes posteriores. Protoarchaeopteryx e Caudipteryx não são pássaros mesmo que tenham penas porque o conjunto de caracteres morfológicos que eles possuem marca-os como pertencentes aos dinossauros coelurosaurianos maniraptóides.]
Parece que as penas não podem mais ser usadas como uma característica única dos pássaros.
Para uma discussão sobre os dinossauros com penas, vá para Jeff Poling's Dinosauria page.
- 2) Hallux oponente (dedão grande).
-
Este também é um caráter dos pássaros e não dos dinossauros. Embora dedões grandes oponentes sejam encontrados em outros grupos, eles não são, quanto ao meu conhecimento, encontrados em dinossauros. Um dedão grande reverso é encontrado em alguns dinossauros, no entanto, e a condição é aproximada em alguns dinossauros terópodes.
- 3) Furcula (osso do peito) formada por duas clavículas fundidas juntas na linha média.
-
Agora começamos a entrar em terreno instável. Antigamente pensava-se que a posse de uma furcula distinguia pássaros de dinossauros. De fato, até recentemente até clavículas eram poucas e distantes em dinossauros terópodes mesmo (o grupo sugerido mais próximo aos pássaros e do qual os pássaros evoluíram - veja Ostrom 1976). No entanto, foi encontrado que os dinossauros terópodes realmente tinham clavículas (por exemplo, Bryant & Russell 1993) e elas foram encontradas em várias espécies, por exemplo, Segisaurus, Velociraptor, Euparkeria, Ornithosuchus, Saltoposuchus, Ticinosuchus. Além disso, Chure & Madson (1996) relataram furculas em um dinossauro alosaúrido não maniraptóide.
Foi encontrado que as clavículas são frequentemente pequenas e pouco ossificadas. Isso não é surpresa, pois elas são de pouca vantagem evolutiva para o dinossauro terópode médio. No entanto, os pássaros também mostram essa variação em ossificação, especialmente entre os carnívoros e alguns papagaios, as clavículas são reduzidas ou até mesmo ausentes. Portanto a aparente ausência de clavículas em alguns dinossauros terópodes pode ser devido à pouca ossificação em vez de ausência verdadeira. No entanto, furculas foram encontradas em alguns dinossauros terópodes, nomeadamente os Oviraptorosauria (Barsbold et al. 1990, Bryant & Russell 1993), por exemplo Oviraptor e Ingenia. Assim as furculas não parecem ser diagnósticas para pássaros e certos membros do grupo sugerido mais próximo aos pássaros agora parecem possuir furculas, então é um caráter neutro.
Uma crítica comumente citada disso é que a maioria dos dinossauros terópodes listados aqui são posteriores ao Archae. No entanto, nenhum deles é reivindicado como o ancestral de qualquer forma, e Eupakeria é uma forma do Triássico. A presença de clavículas mostra que este caráter é uma característica de dinossauros terópodes e, portanto, provavelmente estava presente em terópodes iniciais.
- 4) Púbis alongado e dirigido para trás.
-
Esta é uma característica dos pássaros, mas também é uma característica de alguns dinossauros terópodes, então não é diagnóstica dos pássaros - outro caráter neutro. No entanto, os eixos púbicos de Archaeopteryx e dromaeossaurídeos (um grupo de dinossauros terópodes que são pensados como estando intimamente ligados aos pássaros) compartilham uma seção transversal transversal ligeiramente inclinada em forma de placa que não é encontrada em nenhum outro arcosauriano.
As características reptilianas do Archaeopteryx
- 5) A pré-maxila e a maxila não estão cobertas por chifres.
-
Esta é uma forma elegante de dizer "não possui bico". A premaxila não possui uma cobertura queratinizada, portanto Archaeopteryx não tem bico. O bico é produzido através do processo de 'cornificação', que envolve a camada de muco da epiderme (Romanoff 1960) e, portanto, sua formação é independente da formação do osso da mandíbula.
- 6) As vértebras da região do tronco são livres.
-
Em aves, as vértebras do tronco estão sempre fundidas.
- 7) Os ossos são pneumáticos.
-
Ou seja, parecem ter sacos aéreos, como ocorre em aves e em alguns dinossauros (por exemplo, Witmer 1990, Brooks 1993). Deve-se ressaltar que alegações anteriores sugerindo que os ossos do Archae não eram pneumáticos (Lambrecht 1933; de Beer 1954) basearam-se em evidências negativas, ou seja, que os ossos não exibem poros pneumáticos (através dos quais os sacos aéreos entram nos ossos) e os ossos não apresentam nenhuma das protuberâncias e abaulamentos que caracterizam os ossos pneumáticos das aves modernas. Britt et al. (1998) encontraram evidências da presença de ossos pneumáticos em Archaeopteryx:
"Aqui reexaminamos dois espécimes de _Archaeopteryx_. Estes espécimes mostram evidências de pneumaticidade vertebral nas vértebras cervicais e torácicas anteriores, confirmando assim a continuidade filogenética entre os sistemas pneumáticos de terópodes não-avialanos e as aves vivas" (Britt et al. 1998, p. 374)
8) Shafts púbicos com seção transversal transversal achatada e levemente inclinada-
Uma característica compartilhada com dromaeossáurios, mas não com outros dinossauros ou aves
- 9) Os hemisférios cerebrais alongam-se, tornam-se alongados e o cerebelo situa-se atrás do cérebro médio, não o cobrindo por trás nem pressionando-o para baixo.
-
Esta é novamente uma característica reptiliana. Nos pássaros, os hemisférios cerebrais são robustos, o cerebelo é tão ampliado que se estende para frente sobre o cérebro médio e o comprime para baixo. Assim, a forma do cérebro não é como a dos pássaros modernos, mas sim uma etapa intermediária entre dinossauros e pássaros (por exemplo, Alexander 1990).
- 10) O pescoço se conecta ao crânio pela parte traseira, como nos dinossauros, e não por baixo, como nos pássaros modernos.
- O local de fixação do pescoço (visto de baixo) é característico em aves; o _Archaeopteryx_ não possui essa característica, mas é o mesmo que nos dinossauros terópodes:
"Observe que esse pescoço semelhante ao dos coelurosáurios estendia-se para trás a partir da parte traseira do crânio no _Archaeopteryx_ — como ocorre nos coelurosáurios [dinossauros terópodes] — e não a partir de baixo, como em aves mais recentes." (Ostrom 1976, p. 137).
O crânio e o cérebro do Archae são basicamente reptilianos e não são "totalmente semelhantes aos de aves" (contrário a uma certa alegação criacionista).
- 11) O centro das vértebras cervicais possui superfícies articulares côncavas simples.
-
Isso é o mesmo padrão dos arcosaurianos. Nos pássaros, as vértebras são diferentes; elas possuem uma superfície em forma de sela:
"A característica mais marcante das vértebras são as facetas simples e em forma de disco dos seus corpos, sem qualquer sinal das articulações em forma de sela encontradas em outros pássaros" (de Beer 1954, p. 17).
- 12) Cauda óssea longa com muitas vértebras livres até a ponta (sem pigóstilo).
-
Os pássaros têm uma cauda curta e as vértebras caudais estão fundidas para formar o pigóstilo.
- 13) Os ossos pré-maxilar e maxilar possuem dentes.
-
Nenhuma ave moderna possui dentes (por exemplo, Romanoff 1960; Orr 1966, p. 113). Embriões de aves formam brotos dentários, mas não produzem dentes efetivamente. Algumas aves produzem subsequentemente cristas no bico, mas não há conexão entre elas e os brotos dentários embrionários, uma vez que as cristas também se formam em outras áreas do bico onde nenhum broto dentário se formou previamente. Algumas aves produzem estruturas em forma de gancho que são papilas e parecem estar relacionadas ao processo de queratinização do bico (Romanoff 1960), e não têm nada a ver com dentes. Elas não possuem conexões de vasos sanguíneos ou nervos, nem produzem dentina.
A expressão de brotos dentários no embrião de ave tem uma explicação evolutiva simples, uma vez que sugere que os ancestrais das aves modernas possuíam dentes e que esse caráter foi suprimido nas aves modernas. A presença de brotos dentários nos embriões de organismos que não possuem dentes no adulto é uma dificuldade para anti-evolucionistas, uma vez que por que um caráter seria expresso se nunca for usado no organismo? Alguns pássaros fósseis exibem uma redução no número de ossos que possuem dentes. Tanto Hesperornis quanto Baptornis carecem de dentes na premaxila (Archaeopteryx e dinossauros terópodes possuem dentes tanto na maxila quanto na premaxila). Não apenas isso, Hesperornis possui um bico, mas apenas na mandíbula superior (Gingerich 1975). Portanto, ele tem metade de um bico e dentes. Um bom exemplo de uma estrutura morfologicamente intermediária entre aves dentadas que carecem de bico e aves com bico, sem dentes.
- 14) Costelas finas, sem articulações ou processos unciformes e não se articulam com o esterno.
-
Os pássaros possuem costelas robustas com processos uncinais (suportes entre elas) e articulam-se com o esterno.
- 15) O cinto pélvico e a articulação do fêmur são arcosaurianos, e não avianos (exceto pelo púbis apontando para trás, como mencionado acima).
-
Aqui o Archae realmente mostra sua natureza transicional. Embora a cintura pélvica como um todo seja basicamente livre e semelhante às cinturas de arcosáurios, o púbis aponta para trás — uma característica compartilhada com aves e alguns outros dinossauros terópodes semelhantes a aves.
O que é interessante é que, com a pélvis das aves:
"O ísquio situa-se abaixo da parte posterior do ilíaco e abaixo deste, novamente, está o púbis, que é dirigido para trás (ou seja, assim: =). Estudos embriológicos mostram que a posição peculiar desses ossos é o resultado de uma rotação secundária e que o processo pectíneo, à frente do acetábulo, não é o verdadeiro púbis, como alguns pesquisadores sustentaram." (Bellairs & Jenkin 1960, p. 258).
Em outras palavras, a pélvis embrionária da ave, quando primeiramente formada, parece, em forma e ângulo entre o ilíaco e o púbis (45 graus), muito semelhante à pélvis em formato de "A" do Archaeopteryx (ou seja, assim: <) (por exemplo, Romanoff 1960). A pélvis totalmente formada, com todos os ossos alinhados paralelamente, é o resultado da rotação secundária do púbis de "<" para "=". Isso apoia a visão de que as aves tiveram um ancestral com uma pélvis saurischiana, como o tipo possuído pelo Archaeopteryx e outros dinossauros terópodes. (veja também A história de duas pélvises abaixo)
- 16) O Sacro (as vértebras desenvolvidas para a fixação da cintura pélvica) ocupa 6 vértebras.
-
Isso é o mesmo que em répteis e especialmente em dinossauros ornitípodos. O sacro das aves cobre entre 11-23 vértebras! Portanto, embora a variação observada em aves modernas seja grande, está longe do número encontrado em Archaeopteryx
- 17) Metacarpos (mão) livres (exceto o 3º metacarpo), articulação mão-pulso flexível.
-
Isso é como em répteis. Nos pássaros, os metacarpos estão fundidos com os carpais distais no carpo-metacarpo, pulso/mão fundidos. Todos os pássaros modernos têm um carpo-metacarpo, todos os pássaros fósseis têm um carpo-metacarpo - exceto um (adivinhe!) :-). No entanto, os carpos de vários grupos de dinossauros coelurosaurianos mostram uma tendência para fusão, e na forma do Cretáceo Superior Avimimus, um verdadeiro carpo-metacarpo é formado.
Tem sido sugerido que o avestruz e/ou outros Ratites também possuem ossos de pulso/mão não fundidos. Isso não é correto:
"O avestruz, emus, avestruzes, casuaris e kiwis são frequentemente referidos juntos como os Ratites, embora possam não estar estreitamente relacionados entre si. Eles têm asas pequenas e não podem voar, mas os ossos de suas mãos estão fundidos juntos da mesma maneira peculiar que nos pássaros voadores, o que sugere que evoluíram de pássaros voadores." (Alexander 1990, p. 435).
Alguma semelhança entre a mão do avestruz e alguns dos dinossauros terópodes mais derivados foi uma vez usada para sugerir que os Ratites eram 'primitivos' e evoluíram antes do advento do voo nos pássaros. No entanto, Tucker (1938b) mostrou que tais semelhanças são inteiramente superficiais.
"Ele dirigiu a atenção para os caracteres semelhantes aos de pássaros da mão do dinossauro Ornitholestes como evidência de que uma mão semelhante a de pássaro pode ser desenvolvida independentemente do voo, mas o autor apontou na comunicação mencionada acima [Tucker 1938b] que a semelhança é inteiramente superficial e que a peculiar curvatura e fusão terminal dos metacarpos 2 e 3 que caracterizam tanto a mão Carnate quanto a mão Ratite não são de forma alguma [sic?] reproduzidas no dinossauro." (Tucker 1938a, p. 334).
"Voltando agora às razões nas quais se buscou basear a visão de que os Ratites eram pássaros primitivos cujos ancestrais nunca voaram, um: a semelhança entre a mão do avestruz e a do dinossauro, foi descartada como inválida. Tucker (1938b) mostrou que tais semelhanças que existem entre eles são apenas superficiais e sem significado." (de Beer 1956, p. 65).
- 18) Abertura nasal muito à frente, separada do olho por uma grande fenestra pré-orbital (furo).
-
Isso é típico de répteis, mas não de aves. Quando uma fenestra está presente em aves, ela está sempre muito reduzida e está envolvida na proquinesia (movimento do bico)
- 19) A crista deltoidiana do úmero enfrenta anteriormente, assim como os côndilos radial e ulnar.
-
Típico de répteis, mas não encontrado em aves
- 20) Garras em 3 dígitos não fundidos.
-
Nenhum pássaro adulto moderno tem 3 garras, nem possuem dedos não fundidos. O hoatzin juvenil e os touracos possuem 2 garras, mas perdem-nas conforme crescem; o avestruz parece reter suas 2 garras até a idade adulta, devido ao término precoce do desenvolvimento (veja a seção sobre Ratites). No caso do hoatzin, acredita-se que essas garras permitem que o juvenil suba. Já se alegou que, como esses pássaros possuem garras, mesmo na fase juvenil, a presença de garras não pode ser usada como um caráter reptiliano. Isso não é verdade, no entanto. Na verdade, quase todos os pássaros exibem garras, mas na fase embrionária e são perdidas até que o pássaro saia do ovo. No caso dos poucos que retem garras até a fase juvenil, isso é meramente a extensão da condição para a fase pós-embrionária. Como McGowan (1984, p. 123) diz:
"Ao reter uma característica reptiliana primitiva que outros pássaros perdem pouco antes de sair do ovo [o hoatzin], está nos mostrando sua linhagem reptiliana. Longe de ser evidência em contrário, o hoatzin é evidência adicional para a ancestralidade reptiliana dos pássaros."
- 21) A fíbula é igual em comprimento à tíbia na perna.
-
Isso novamente é uma característica típica dos répteis. Nos pássaros, a fíbula é encurtada e reduzida.
- 22) Metatarsos (ossos do pé) livres.
-
Em aves, estas são fundidas para formar o tarsometatarso. No entanto, em embriões de aves modernas, os ossos do pé são inicialmente separados, como no adulto Archaeopteryx e é outro caráter que apoia uma ancestralidade reptiliana para as aves. Afinal, por que se preocupar em produzir ossos separados no embrião e depois fundi-los? Por que não produzir uma massa fundida para começar? Nenhum ave moderna adulta possui metatarsais separados, mas eles estão separados, inicialmente, no embrião. Isso pode ser explicado em termos de evolução - as aves evoluíram de um grupo que possuía metatarsais não fundidos.
Ceratossauros, Avimimus e Elmisauridae todos apresentam verdadeiros tarso-metatarsos. O próprio Archae mostra apenas o início desta estrutura.
- 23) Presença de gastrálias.
-
Gastrálias são "costelas ventrais", elementos de osso dérmico na parede ventral do abdômen. Típicas de répteis, elas estão ausentes em aves, por exemplo:
"Além das verdadeiras costelas, o espécime do British Museum mostra um grande número de assim chamadas costelas ventrais ou gastrálias, elementos de osso dérmico situados na parede ventral do abdômen." (de Beer 1954, p. 18)
"As gastrálias do espécime de Berlim são idênticas às do espécime do British Museum, mas mais foram preservadas." (de Beer 1954, p. 19)
"O 'novo' espécime foi encontrado em 8 de setembro de 1970 exposto no Museu Teyler, Haarlem, Países Baixos. Ele consiste em duas pequenas lajes (espécimes 6928 & 6929), parte e contraparte que contêm impressões ou partes da mão esquerda e antebraço, pélvis, ambas as pernas e pés, e algumas gastrálias." (Ostrom 1970, p. 538)
"Também presentes são numerosos fragmentos de gastrálias, impressões fracas de três ou quatro vértebras dorsais, . . " (Ostrom 1972, p. 291).
"A laje contraparte (No. 6929) contém gastrálias adicionais, falanges, .." (Ostrom 1972, p. 291)
"Gastrálias, ou costelas abdominais dérmicas, estão presentes em todos os cinco espécimes esqueléticos de _Archaeopteryx_" (Ostrom 1976, p. 139-140).
Gastrálias estão presentes no espécime de Eichstatt (Veja Wellnhofer 1974, fig. 7C)
Tabela de características de Archaeopteryx
1 = presente; * = presente em alguns; ? = possivelmente presente; x = ausente
Dinossauros Archae Aves
1 * 1 1
2 x 1 1
3 * 1 1
4 * 1 1
5 x x 1
6 x x 1
7 * x 1
8 * 1 x
9 1 1 x
10 1 1 x
11 1 1 x
12 1 1 x
13 1 1 x
14 1 1 x
15 1 1 x
16 6 6 11-23
17 1 1 x
18 1 1 *
19 1 1 x
20 1 1 x
21 1 1 x
22 1 1 x
23 1 1 x
Pode-se ver que o Archae possui muitos mais caracteres que estão presentes em dinossauros e não em pássaros, do que caracteres que estão presentes em pássaros mas não em dinossauros. É por isso que o Archae é uma verdadeira espécie transicional, porque compartilha alguns caracteres que são diagnósticos de um grupo, enquanto ainda mantém caracteres diagnósticos do seu grupo ancestral. Qualquer um que afirme que o Archae é 100% pássaro está errado. Qualquer um que afirme que o esqueleto do Archae é até predominantemente parecido com o de pássaros está errado. Qualquer um que afirme que o Archae tem um crânio "totalmente parecido com o de pássaro" está errado.
Este último ponto é feito em referência à alegação do Dr. Duane Gish de que o crânio de Archae é "totalmente parecido com o de um pássaro" (R. Trott, comunicação pessoal, 1994). Esta alegação é falsa. Para demonstrar isso, precisamos considerar o crânio de Archae com mais atenção.
Características cranianas de Archaeopteryx
Como mencionado acima, o Dr. Gish afirma que o crânio de Archae é "totalmente semelhante ao de um pássaro". Isso é falso. Romer (1950 p. 261) descreve Archae da seguinte forma: "O crânio, na medida em que pode ser visto, era bastante semelhante ao de um pássaro...". No entanto, não apenas isso está muito longe de "totalmente semelhante ao de um pássaro", mas Romer estava utilizando a reconstrução detalhada do Especime de Berlim, por Heilmann (1926). Ostrom (1976 p. 131) diz o seguinte sobre a reconstrução de Heilmann:
"Apesar dos detalhes mostrados lá [reconstrução do crânio de Heilmann -cn], o espécime real não permite conclusões tão detalhadas e precisas. O crânio do espécime de Berlim - cn está muito amassado e os ossos estão extensivamente fraturados, lascados e distorcidos - a ponto de que muito poucas suturas cranianas ou mandibulares são inequivocamente identificáveis. As reconstruções de Heilmann foram republicadas por muitos autores e interpretações e hipóteses subsequentes baseadas nelas. Provavelmente, alguns autores não estavam cientes da base inadequada da reconstrução de Heilmann, e compreensivelmente, a menos que tenham tido a oportunidade de examinar o próprio espécime."
E novamente na mesma página:
"Felizmente, o espécime de Eichstatt agora fornece uma base comparativa para avaliar e corrigir reconstruções passadas do crânio de Berlim."
Como mencionado acima, o espécime de Eichstatt não foi descrito até 1974, portanto a descrição de Romer baseou-se na reconstrução de Heilmann.
Usando ambos os espécimes, Ostrom (1976 p. 132) delineou 10 caracteres no crânio de Archae que ele compartilha com outros dinossauros terópodes, como Ornitholestes, Compsognathus, Velociraptor e talvez Saurornithoides. Estes são:
- Um bico afilado e pontiagudo. Nos pássaros, a maxila forma o "bico". Embora o bico esteja encurtado em Archae.
- Nares externas longas e elípticas [narinas] delimitadas quase exclusivamente acima e abaixo pela premaxila e nasal (um osso na parte frontal da mandíbula superior). Nos pássaros, as narinas são reduzidas e removidas posteriormente (mais próximas dos olhos) devido ao aumento do tamanho da premaxila.
- Uma grande fossa pré-orbital e uma grande fossa pós-orbital triangular que contém duas pequenas aberturas. (Estes são orifícios no crânio. Um é uma única abertura à frente da órbita ou cavidade ocular, o outro está atrás da órbita e possui duas aberturas). Em outros pássaros fósseis, a pré-orbital está reduzida, a pós-orbital possui apenas uma abertura. Nos pássaros modernos, a pré-orbital está muito reduzida e a pós-orbital possui uma única abertura.
- Uma barra pré-orbital fina e quase vertical separando a fossa antorbital e a órbita (Aqui está o buraco novamente! Isso significa que há uma barra vertical de osso separando o buraco à frente do olho do olho em si). Pássaros fósseis e modernos possuem uma barra pré-orbital muito mais robusta - devido à redução da fossa pré-orbital.
- Uma órbita grande e circular que contém um grande an esclerótico. (O an esclerótico é comum a répteis, pássaros e peixes actinoptígeos, mas a maioria dos répteis fósseis e todos os pássaros fósseis possuem. São uma série de placas ossificadas que circundam o olho).
- Um osso jugal fino e reto forma o arco zigomático (este é o osso que corre abaixo do olho, a maçã do rosto conforme se estende para trás em direção à orelha, nos humanos). Nos pássaros modernos, o arco é composto pelos ossos quadratojugal, jugal e maxila.
- Um quadrato robusto de comprimento moderado que está inclinado para frente. Este é o osso na mandíbula superior que forma parte da articulação da mandíbula - com o articular - nos répteis, e o Archae possui um grande, hum, se você entende o que eu quero dizer! (A propósito - para aqueles de vocês que ainda estão conosco, o quadrato está conectado ao estapés na mandíbula superior, e como todos sabemos, o estapés é o osso que vibra na orelha para que possamos todos ouvir. Assim, o estapés e o quadrato estavam conectados nos répteis e não é um grande salto para ter tanto o estapés quanto o quadrato na orelha como ocorre nos mamíferos. Assim, a alegação feita por Gish de que - para que os ossos entrassem na orelha na transição de répteis para mamíferos, eles teriam tido que passar por uma fase na qual o 'mamífero' teria surdo a cada vez que abrisse a boca - não é precisa, já que a condição de articulação do estapés+quadrato na articulação da mandíbula é encontrada em répteis fósseis e extantes, mas eu me alongo).
- Uma mandíbula inferior que é incomumente rasa e possui uma curva notável atrás da fileira de dentes. Não nos pássaros.
- Um processo retroarticular longo. Novamente, uma característica clássica da articulação da mandíbula de répteis, mas compartilhada com os pássaros.
- Os ossos premaxila, lacrimal e jugal estão separados. Nos pássaros, o osso premaxila está conectado ao osso jugal; o osso jugal está conectado ao osso lacrimal. . .
:-)
- Um único vomer.
- Curvatura forte do gancho ectopterigóide.
- Um recesso dorsal no proótico.
Embora Archaeopteryx possua certos caracteres cranianos que são avianos, estes são insuficientes para permitir a caracterização "totalmente semelhante a um pássaro".
Heilmann descreveu uma fenestra mandibular externa (um orifício na mandíbula inferior) delimitada abaixo pelo osso dentário — que é a condição encontrada nos pássaros — e não pelo osso angular — como ocorre em répteis. Isso indicaria que o Archae possui uma mandíbula inferior que continha tanto características avianas (fenestra mandibular mencionada) quanto características reptilianas (mandíbula inferior dentada). Tudo isso está muito bem e é algo que se gostaria de ver no Archae. No entanto, como Ostrom (1976, p. 132) aponta:
"Embora eu gostaria muito de aceitar essa interpretação, o estado altamente fragmentado do osso da mandíbula inferior (ou ossos) que margeia a suposta fenestra mandibular, seja abaixo ou acima, torna impossível certificar suas identificações. De fato, as margens superiores fragmentadas da suposta fenestra deixam dúvidas consideráveis quanto à própria existência de uma 'fenestra' - uma dúvida que não foi removida pelo espécime de Eichstätt." Assim, longe de conspirar para apresentar o Archae na melhor luz possível, uma característica muito útil que teria auxiliado na descrição do Archae como um verdadeiro fóssil transicional mostrou-se, na verdade, em toda probabilidade, não verdadeira. Tanto faz com a conspiração evolucionista! De fato, o sétimo espécime mostra que uma fenestra mandibular está ausente (Elzanowski & Wellnhofer 1996)
Outra característica importante do crânio de Archae é o côndilo occipital e o forame magno. Em Archae, estes estão bem acima da extremidade dorsal do quadrado. Como Ostrom (1976 p. 136) escreve:
"Esta condição primitiva é característica tanto dos pseudossucianos quanto dos terópodes, em contraste com todos os pássaros posteriores onde o côndilo occipital e o forame magno estão na base do crânio, bem abaixo do nível da extremidade superior do quadrate. Neste aspecto, Archaeopteryx estava longe de ser aviano."Whetstone (1983, p. 449) does describe the caixa craniana (as opposed to the cabeça) of Archaeopteryx as, "typically avian", however, he also describes skull features found in Archaeopyteryx and not in birds.
Recentemente, surgiram algumas especulações sobre a estrutura do osso quadrate em Archae. O quadrate do espécime de Eichstatt foi descrito como de cabeça única - ou seja, o topo do quadrate possui apenas um botão arredondado que se articula com o crânio em apenas um local. Em todas as aves modernas, o quadrate é de cabeça dupla - ou seja, o topo do quadrate possui dois botões arredondados e, portanto, se articula com o crânio em dois locais. Com base na tomografia computadorizada de uma porção enterrada do crânio do espécime de Eichstatt, Haubiltz et al. (1988) sugeriram que Archae possuía um quadrate de cabeça dupla. No entanto, a imagem é de baixa qualidade e não se pode descartar a presença de outro osso subjacente ao quadrate. O único quadrate encontrado in situ em qualquer espécime é de cabeça única e um osso similar do espécime de Londres, que parece ser o quadrate, também é de cabeça única. A identificação do quadrate no sétimo espécime confirmou que o osso era de cabeça única (Elzanowski & Wellnhofer 1996). Assim, o quadrate de Archae parece ser reptiliano e não aviano.
Como se pode ver, o crânio do Archae não é "totalmente aviar". A alegação de ser "totalmente aviar" carece de fundamento.
Uma é que as diferenças observadas entre _Archaeopteryx_, aves fósseis e aves modernas podem ser explicadas como apenas variação dentro das aves. Associada a isso está a alegação de que algumas aves modernas (Ratitas) compartilham um número de características morfológicas com _Archaeopteryx_.
Archaeopteryx + aves modernas, apenas variação dentro da espécie?
Foi sugerido que as diferenças entre Archaeopteryx e as aves modernas representam uma simples variação intra-grupal. No entanto, isso não é correto. As aves modernas apresentam um grande número de caracteres morfológicos derivados não possuídos por Archaeopteryx. Morfologicamente, Archaeopteryx claramente parece estar mais estreitamente relacionado aos dinossauros terópodes do que a qualquer outro grupo e é agrupada com as aves em vez dos dinossauros terópodes devido à posse de apenas dois caracteres principais: presença de penas e presença de um hálux (dedo) totalmente revertido.
A variação morfológica relevante pode ser representada graficamente (de forma grosseira) na figura a seguir:
|
^ | A = Archaeopteryx
| |
| ___________________
T | | |
i | | PÁSSEROS MODERNOS |
m | __ __ __ __ __| |
e | | |___________________|
| | PÁSSEROS FOSSÍIS |
| __| BIRDS |
| | A|__ __ __ __ __ __|
| |___|
|______________________________________________
"réptil" "pássaro"
<--- Morfologia -->
Como pode ser visto, a variação dentro das aves mostra uma tendência distinta. As morfologias mais semelhantes a "répteis" ocorrem nas aves mais antigas, com a morfologia típica da ave "moderna" restrita às aves mais recentes. Se, como sugerido, a variação morfológica é simplesmente variação dentro das aves, esperaríamos ver os vários grupos morfológicos (aves fósseis, aves modernas) distribuídos uniformemente ao longo do intervalo de tempo relevante. Se, no entanto, as aves evoluíram de dinossauros terópodes, então esperaríamos ver as primeiras aves possuírem mais caracteres semelhantes a "répteis" e as aves modernas mais derivadas possuírem menos caracteres semelhantes a "répteis". Isto, de facto, é o que observamos. Assim, a distribuição de caracteres dentro das aves apoia a sua origem a partir de ancestrais dinossauros terópodes e não apoia a alegação de que a variação é simplesmente 'dentro da espécie'.
Ratítes
Muita ênfase é dada às características morfológicas dos Ratitas (avestruzes, kiwis, etc.), e superficialmente pode parecer que essas aves apoiam a sugestão de 'variação dentro da espécie', conforme mostra a figura a seguir:
_____________________________
| | |
|RATÍTES| PÁSSEROS MODERNOS |
|_______| |
|_____________________|
________________________________________________
"réptil" "pássaro"
<--- Morfologia -->
Importante, no entanto, outros caracteres morfológicos possuídos pelos ratitas mostram claramente que eles são derivados de uma morfologia de aves modernas, voadoras e, portanto, os ratitas são na verdade neotônicos em relação às aves modernas, com seus caracteres mais semelhantes a "répteis" devido ao término precoce do desenvolvimento, deixando-os com alguns caracteres morfológicos semelhantes ao Archaeopteryx e a algumas aves fósseis (veja abaixo). Esses caracteres são de desenvolvimento e, portanto, fornecem evidências adicionais de suporte para uma ligação entre dinossauros terópodes e aves.
Uma história de dois pélvis
Os dinossauros podem ser divididos em dois grupos com base na forma do quadril. Os Saurischia, ou dinossauros de "quadril de lagarto", e os Ornithischia, ou dinossauros de "quadril de ave". Estranho como possa parecer, acredita-se que o Archaeopteryx e todas as aves modernas tenham evoluído dos Saurischia, e não dos Ornithischia, de "quadril de ave"! Estranho? Bem, não exatamente. Os quadris em questão são mostrados na Fig.1.
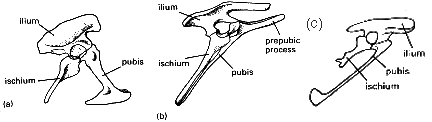
O pélvis saurísquio (Fig. 1a) é a estrutura mais primitiva, e tem o púbis apontando para frente e o ísquio para trás. Nos ornitísquios (Fig. 1b), o púbis corre para trás em paralelo com o ísquio. Além disso, está presente um processo pré-púbico, que é assumido ter sido usado como ponto de fixação muscular na ausência do púbis (provavelmente para suportar o intestino). Esta situação é superficialmente semelhante àquela em aves (daí o nome 'de anca de ave' - o pélvis das aves era bem conhecido pelos anatomistas até o momento em que os dinossauros estavam sendo descritos). No entanto, em um exame mais detalhado, diferenças claras são evidentes. Em Archaeopteryx (Fig. 1c) e todas as aves, o processo pré-púbico está ausente e, ao contrário dos saurísquios, os púbis não se encontram ao longo da linha média. Além disso, em Archaeopteryx o púbis ainda mantém a terminação em forma de clava, como em muitos saurísquios. Também deve ser notado que vários dinossauros saurísquios têm o púbis voltado para trás, por exemplo, Troodon. O pélvis de Archaeopteryx é morfologicamente intermediário entre dinossauros saurísquios e aves.
Mas por que isso ocorreu? Bem, o principal gerador de impulso em terópodes anteriores era o Caudofemoralis longus, que se estendia da perna até uma certa distância ao longo da cauda. Formas posteriores, como os dromaeossáuridos (o grupo estreitamente relacionado ao Archaeopteryx), reorganizaram isso de modo que o impulso primário fosse produzido por músculos que se ligavam da perna ao pélvis e à base da cauda. Isso permitiu que a cauda fosse estabilizada e usada como um dispositivo de estabilização, como nos alossauros e nos "raptors", ou que fosse perdida, como nas aves modernas.
Em ornitísios, a pubis apontando para trás provavelmente ocorreu para permitir a expansão do estômago e do intestino a fim de digerir melhor o material vegetal.
O estudo do desenvolvimento embrionário do pélvis das aves modernas também é instrutivo. O pélvis embrionário nas aves modernas começa em uma configuração muito semelhante à de Archaeopteryx, com os ossos ilíaco e púbico posicionados separadamente a um ângulo de aproximadamente 45 graus. Os ossos embrionários então giram secundariamente por ângulos observados em modernas Ratites até alcançarem o alinhamento paralelo que caracteriza as aves adultas e modernas (ver Romanoff 1960, p. 1010):
Embrião de galinha Ângulo ílio-púbis Adultos com o mesmo ângulo
7.0 dias 45 graus Archaeopteryx
7.3 dias 35 graus Kiwi
8.0 dias 30 graus Avestruz
Eclosão <5 graus Galinha moderna
O ângulo entre o ílio e os ossos púbicos na bacia dos ratitas modernos é intermediário entre o Archaeopteryx e os pássaros modernos, como seria de esperar se:
A) Pássaros modernos descendem de ancestrais semelhantes a répteis, similares ao Archaeopteryx
B) Os ratitas são descendentes neotênicos de aves voadoras, nos quais o desenvolvimento foi interrompido prematuramente.
Volar ou não voar
Volar é uma atividade complicada. No entanto, o voo confere uma vantagem evolutiva tão forte que não é surpreendente que essa capacidade tenha evoluído várias vezes. O sucesso dos pássaros fornece ampla evidência para os benefícios positivos do voo. Se o Archae voasse, ele deve ter utilizado muito o mesmo método que os pássaros utilizam hoje, portanto, uma discussão sobre a possibilidade de voo no Archae deve considerar aquelas estruturas mais relevantes para o voo nos pássaros. Estas são as penas, a flexibilidade da asa, a massa muscular e a presença de um esterno cunhado.
Penas
As penas são compostas por um raque central longo e afilado, que suporta ramos laterais espaçados de forma próxima, chamados de barbas. As barbas em ambos os lados do raque constituem uma superfície chamada vane. As duas vanes da pena podem ser simétricas (ou seja, da mesma largura) ou assimétricas, neste caso o raque parece estar mais próximo de uma das bordas da pena do que da outra. As penas de voo dos pássaros modernos são tipicamente assimétricas, enquanto as penas de contorno corporal e semiplumas são simétricas.
Em aves modernas, as remiges, ou penas das asas, são altamente modificadas para o voo de potência (por exemplo, McFarland et al. 1985), principalmente porque o raque é deslocado em direção à borda de ataque da pena (ou seja, a vane de ataque é mais fina que a vane de reboque), resultando em uma pena assimétrica. A vane mais fina ou de ataque da pena sobrepõe a vane mais larga ou de reboque da pena à sua frente (Fig. 2). A vane de reboque contém zonas de barbulas de fricção que seguram a pena sobreposta e impedem que as penas deslizem umas das outras com demasiada facilidade.
Fig. 2
* * * O * * * * *
* * * O * * * * *
* * * O * * * * *
| | |
| | |--pavão traseiro
| |---------eixo
|-----------pavão dianteiro
Speakman & Thomas (1994) compararam a assimetria de algumas das penas de voo de Archae (penas de voo 4, 5 e 6) com as de aves modernas voadoras e não voadoras e com a pena isolada de Solnhofen. Eles descobriram que a assimetria média para as penas de Archae foi de 1,25, o que é menor do que o das aves voadoras modernas (o mínimo em torno de 2,2), mas que se sobrepõe àquela das aves modernas não voadoras. A pena isolada exibiu uma assimetria de 2,2 - apenas dentro da faixa das aves voadoras modernas. No entanto, não se sabe se esta pena é de Archae, ou onde na Archae a pena estava situada, caso seja uma pena de Archae. Norberg (1995) corretamente apontou que a curvatura da pena também é importante para a capacidade de voo, e as penas de Archie exibem curvatura significativa. Parece que Speakman & Thomas podem ter medido as penas erradas, no entanto. Paul Davis (com. pers. 1996) apontou que Speakman & Thomas parecem ter numerado as penas primárias começando na ponta da asa e movendo-se em direção ao corpo, enquanto a prática atual é numerar as penas começando do corpo e movendo-se para fora ao longo da asa (por exemplo, Rietschel 1985). Isso é importante, já que a assimetria das penas diminui em direção à ponta da asa - assim Speakman & Thomas parecem ter medido as penas com menor assimetria. Medições das penas de maior assimetria próximas ao corpo (boby) resultam em valores de cerca de 2,32 - o que se sobrepõe à faixa inferior para aves voadoras existentes (por exemplo, Norberg 1995; P.G. Davis com. pers. 1996). As primeiras penas de voo das aves são sempre altamente assimétricas, no entanto, e por isso este resultado é ambíguo. A maioria das penas de voo de Archae parece ter uma ligeiramente menor assimetria do que aquela encontrada em aves voadoras hoje. Na realidade, no entanto, a assimetria da pena é um pouco de um cavalo de Troia quando se trata de capacidade de voo. As penas são assimétricas para manter a integridade aerodinâmica à medida que se deformam sob cargas aerodinâmicas durante o voo, e assim a maior assimetria ocorre em aves nas quais tais cargas são maiores - ou seja, aves com alta manobrabilidade e voo de subida (J.M.V. Rayner, com. pers. 1995). Assim, a assimetria é uma condição derivada para uma capacidade de voo derivada - uma Archaeopteryx nunca possuía. Assim, não é surpreendente que Archaeopteryx possuísse penas de baixa assimetria, mas isso não impede Archaeopteryx de voar.
Flexibilidade das asas
Considere a asa dos carniáves [pássaros com esterno em forma de quilha]. Uma maravilha da adaptação, perfeitamente capaz de sustentar animais bastante grandes em voo de força por longas distâncias. Mas o que é que na anatomia da asa permite a possibilidade de voo? Bem, simplesmente dito, 'está tudo na ação do pulso'.
Na aves modernas, a articulação do ombro atua como uma articulação universal, com um alto grau de mobilidade não apenas em adução [movimento da asa para baixo], abdução [movimento da asa para cima] e extensão [estender a asa afastando do corpo] e retração [puxar a asa para dentro do corpo], mas também em rotação significativa ao longo do eixo longitudinal do úmero.
O cotovelo, por contraste, é uma articulação muito restritiva. Ele permite apenas extensão e flexão planar [ou seja, desdobramento e dobramento da asa - ou seja, não há componente rotacional].
No entanto, é a articulação do pulso que é importante. O pulso é composto por dois ossos carpais que controlam o movimento da articulação. Distalmente além disso está o carpometa-carpo fundido – produzido a partir da fusão dos ossos carpais [do pulso] remanescentes e dos metacarpais [da mão inferior]. Além disso está a mão, por vezes incompletamente fundida, composta por três dígitos. Este complexo, distal da articulação do pulso, compreende uma plataforma quase rígida para a fixação das primárias remiges [penas de voo], enquanto as secundárias remiges se fixam ao antebraço.
A configuração da articulação do pulso, que se articula como faz sobre dois pequenos carpais redondos, permite uma gama incrível de movimentos. Para demonstrar isso, você precisa estar sentado confortavelmente no terminal do computador. Estenda os braços como se fosse para digitar (palmas para baixo, partes peludas para cima). Agora, gire as mãos de modo que as palmas fiquem voltadas para cima (como se estivesse carregando algo). Esta rotação é realizada pela articulação do pulso nos pássaros. Observe que, nos humanos, esta ação é realizada pela rotação do antebraço e não do pulso. Agora, retorne as mãos à posição de partes peludas para cima e gire as mãos de modo que a palma fique voltada para o computador (o sinal universal de "pare") e gire a mão para baixo até que as partes peludas apontem para o computador. Esta rotação é realizada pela articulação do pulso nos pássaros, como é nos humanos.
Como você agora demonstrou, o pulso carniano é muito mais flexível do que o pulso humano (você também demonstrou a qualquer colega de trabalho observando que você está insano e em necessidade urgente de férias!). Essa flexibilidade é vital para o voo motorizado, pois permite que o pulso/mão descreva uma figura-8 preguiçosa durante um batimento completo. Lembre-se de que a articulação do cotovelo não permite rotação, mas a asa deve ser capaz de apresentar uma asa aberta sólida no batimento descendente (de potência), mas deve ser capaz de rotacionar a asa durante o batimento de recuperação (ascendente) para minimizar a resistência. Os pássaros realizam isso rotacionando o pulso e o ombro.
No topo do golpe de potência, o pulso está orientado para apresentar a área superficial máxima ao ar (palma voltada para baixo). Durante o golpe de potência, o pulso e a mão descrevem uma "S", ou dois arcos representando o lado esquerdo do laço superior e o lado direito do laço inferior de um "8". No fundo do golpe de potência, o pulso e a mão estão no fundo do "8". Durante o início do golpe de recuperação, o pulso se reorienta 90 graus, apresentando assim a área superficial mínima ao ar. A asa então completa o "8" descrevendo o lado esquerdo do laço inferior e o lado direito do laço superior. É mais fácil executar do que ler. Segure seu braço direito horizontalmente a partir do ombro, com a palma voltada para baixo. Trave o cotovelo para que você não possa dobrar o braço, apenas rotacioná-lo. Agora, o golpe de potência tem formato de "S", mantendo a palma voltada para baixo e o cotovelo travado. No fundo da "S", rotacione a palma 90 graus até que ela aponte para frente (na verdade, você terá que rotacionar todo o braço para fazer isso; nos pássaros, é feito pela articulação do pulso). Agora, o golpe de recuperação é uma "S" espelhada, completando um "8"; conforme você atinge o topo do golpe, a palma deve torcer de volta para apontar para baixo novamente e você estará pronto para o golpe de potência. Repita algumas vezes até pegar a sensação ou até que alguém chame os caras de jaleco branco. É assim que os pássaros voam e, esperamos, você agora possa ver a importância de ter uma articulação do pulso flexível.
Um último ponto importante neste trecho. A dobra da asa ocorre pela contração do M. biceps brachii, que está ligado à ulna. Este movimento dobra automaticamente o metacarpo e a mão para baixo e para dentro, de modo que a dobra na articulação do pulso envolve o M. biceps brachii.
Agora, o ponto de tudo isso é: essa disposição ocorre em Archae? Essa disposição ocorre em terópodes maniraptóranos? Se sim, que uso possível essa articulação do pulso específica poderia ter para terópodes maniraptóranos? Poderia este ser um exemplo de preadaptação (suspiro! Desculpe usar a "P" palavra).
Primeiro, a articulação do pulso de Archae de fato exibe algumas (mas não todas) das características observadas na articulação do pulso carnívoro moderno. Mais notavelmente, um carpo lunar, de forma hemisférica, cuja porção plana articulava-se com o carpometa-carpo, deixando a extremidade arredondada para articular com a articulação do pulso. Isso proporciona uma articulação do pulso flexível, embora não tão flexível quanto na carnívora moderna.
Em segundo lugar, esta configuração é encontrada em vários táxons de terópodes, como Deinonychus, Velociraptor e Stenonychosaurus. Existem dois problemas aqui: primeiro, todas essas formas são posteriores ao período Arqueano; são todas formas do Cretáceo; em segundo lugar, qual seria a utilidade de um pulso ágil e flexível para um grupo de terópodes que são apenas dentes e unhas?
OK, o primeiro problema é fácil de resolver. Era um problema que essa característica ocorreu em terópodes do Cretáceo e em Archae do Jurássico Superior, pois, para ser uma característica importante e demonstrar uma ligação próxima entre terópodes e Archae, os terópodes coevos com os Archae deviam ter tido esse arranjo particular. Introduzindo Coelurus fragilis, um terópode do Jurássico Superior da América. Essa forma possui, de fato, uma articulação do pulso muito semelhante (Ostrom 1980), fortalecendo não apenas a ligação entre terópodes e aves, mas também a ligação entre terópodes-Arche-aves.
O segundo problema é um pouco mais difícil. Por que uma articulação do pulso tão especializada seria mantida em terópodes maniraptóides, que são tão focados em dentes e unhas ao se tratar de matar? Ostrom (1995) sugere que a flexibilidade seria útil onde uma ação de preensão das mãos pudesse complementar a ação de matar dos dentes e/ou unhas. Eu especularei um pouco mais. Em aves modernas, os bíceps estão envolvidos no dobramento da asa. Se isso fosse estendido aos terópodes, a ação de puxar a presa para dentro do corpo tenderia a fortalecer a preensão do terópode, colocando em jogo o maior músculo do braço - o bíceps - em vez de apenas os músculos do pulso. Tal cenário proporcionaria uma vantagem distinta e, portanto, a retenção da articulação do pulso nos terópodes não é surpreendente. O título da sequência de "Jurassic Park" é, portanto, fácil: "Jurassic Park II: O abraço dos Coelossauros"!
Assim, parece que a articulação flexível do pulso pode ter aparecido primeiro nos dinossauros terópodes como uma ajuda para agarrar e reter a presa e foi secundariamente cooptada pelos pássaros como um mecanismo que permitiu o voo. É muito provável que não apenas os dinossauros terópodes fossem os ancestrais dos pássaros, mas que sejam o único grupo que pudesse ser o ancestral dos pássaros modernos.
No entanto, o pulso de Archae não parece ter a flexibilidade necessária para permitir o voo com potência.
Junção do Ombro
A articulação do ombro é também importante para as aves modernas, porque, não só permite a adução completa (movimento da asa para baixo - Fig. 3b), mas também a abdução completa (movimento da asa para cima - Fig. 3d)
Fig. 3 (vista de frente)
Adução Abdução
a) -----( )----- c) -----( )-----
b) /( )\ d) | |
| | | |
| | \( )/
ou seja, a asa pode ser dobrada para cima sobre as costas (experimente e veja o quão difícil é). Essa amplitude total de movimento é importante para permitir que os pássaros modernos voem com potência. Acontece que a articulação do ombro de Archae parece ser intermediária em orientação entre alguns coelurosaurianos (dinossauros terópodes, por exemplo, Deinonychus) e pássaros (Jenkins 1993) - exatamente como se esperaria de uma forma transicional. No entanto, embora a articulação do ombro proporcionasse um grau substancial de elevação do braço (asa), ela não é suficiente para permitir abdução completa como nos pássaros modernos. Assim, a articulação do ombro de Archae não parece permitir a amplitude total de movimento necessária para o voo com potência.
Massa muscular
Em aves modernas, os músculos de voo (o pectoralis) compreendem até 35% do peso do animal (pense em toda aquela carne suculenta na parte frontal de um frango assado ou peru). É essa massa muscular massiva que fornece a energia necessária para o voo batido e a capacidade de decolar a partir de uma posição em pé. Ruben (1991) utilizou uma estimativa de massa muscular pectoralis para o Archae de aproximadamente 9% do peso corporal. Isso é muito menor do que em aves modernas, mas assumiu que o músculo estava anexado à parede torácica cartilaginosa e não a uma quilha - já que uma quilha é desconhecida no Archae. A quilha é uma extensão óssea do esterno para a fixação de excesso de músculo de voo em aves modernas. Se os músculos estivessem anexados à parede torácica no Archae, o comprimento dos músculos pectoralis seria insuficiente para permitir a abdução completa (veja "articulação do ombro" acima). No entanto, a presença de uma pequena quilha cartilaginosa (e, portanto, não preservada) não está fora de questão. (Espécimes de Archie da Pedra Calcária de Solnhofen não mostram evidências de um esterno. No entanto, descobertas recentes de formas semelhantes a Archie da China sugerem que o Archae de fato possuía um esterno (J.M.V. Rayner, Universidade de Bristol, pers.comm.). Portanto, o valor de 9% é um valor mínimo. Este valor está bem abaixo do observado em aves modernas. Rubin sugeriu que a potência máxima de saída dos músculos locomotores de répteis ativos extantes é o dobro da de aves e mamíferos, indicando que, se o Archae possuísse uma fisiologia de réptil, o voo batido era uma possibilidade, mesmo com tal massa muscular pequena. Speakman (1993) questionou isso, indicando que a potência de saída de répteis sugerida por Ruben foi exagerada, e que o Archae não teria tido a potência de saída para sustentar o voo batido. Além disso, Speakman apontou que o limite teórico inferior de massa muscular pectoralis necessária para decolar a partir de uma posição em pé era aproximadamente 16% do peso corporal. Assim, o Archae não era capaz de voo batido ou decolagem a partir de uma posição em pé.
O supracoracoideus é um músculo importante em aves modernas porque é este músculo que eleva ou abduz a asa durante o voo de potência. Archae não possuía a disposição do supracoracoideus encontrada em aves modernas (Ostrom 1974). Assim, Archae não parece ter tido a massa muscular nem a disposição muscular para permitir o voo de potência, nem ser capaz de decolar a partir de uma posição em pé.
Parece que a estrutura e a fisiologia de Archae seriam contrárias ao voo com propulsão. Mas, será que isso é o fim das habilidades acrobáticas de Archae? Na verdade, não. Observe o uso da expressão "voo com propulsão" na discussão anterior. Quase todas as aves modernas capazes de voo com propulsão são aptas para esse tipo de voo. O voo com propulsão é um voo batido controlado em baixas velocidades e a capacidade de decolar a partir de uma posição parada. Este tipo de voo requer uma grande quantidade de massa muscular e é energeticamente muito dispendioso. Portanto, as aves modernas possuem uma massa muscular peitoral que compreende aproximadamente 20-35% da massa corporal. Para acomodar essa massa extra, desenvolveu-se um quelo no esterno. Em outros desenvolvimentos evolutivos, as penas de voo tornaram-se mais assimétricas para auxiliar no voo com propulsão, e o sistema muscular supracoracoideo assumiu a função de auxiliar na elevação ou abdução da asa, já que isso é difícil em baixas velocidades de voo.
O voo com propulsão é para o voo o que a milha em 4 minutos é para a corrida. No entanto, assim como a incapacidade de correr uma milha em 4 minutos não indica necessariamente que alguém é incapaz de correr, a incapacidade de voar com propulsão não indica necessariamente que o Archae era incapaz de voar. Os caracteres relacionados ao voo que vemos nos pássaros modernos são derivados, uma adaptação ao voo com propulsão, e, portanto, sua ausência (ou subdesenvolvimento) no Archae não pode ser usada para sugerir uma incapacidade de voar.
A estrutura da asa de Archae sugere que era incapaz de voo batido com potência em baixas velocidades. No entanto, a quantidade de adução e abdução, juntamente com a maior flexibilidade do pulso em comparação com dinossauros terópodes, sugere que algum tipo de batida era possível, suficiente para manter Archae no ar em velocidades de cruzeiro normais (aprox. 7-9 ms^-1). Embora a baixa massa muscular do peitoral pareça excluir o voo com potência e o decolagem a partir de uma posição parada, deve-se notar que algumas aves voadoras possuem uma massa muscular inferior ao limite inferior de 16% do peso. Estas incluem alguns mergulhões, que compensam a falta de massa muscular tendo uma decolagem com corrida longa (Speakman 1993). Archae está bem equipado com um aparelho de locomoção terrestre. Assim, é possível que Archae pudesse contornar este problema da mesma maneira que os mergulhões.
Volar a velocidades de cruzeiro normais requer muito menos energia do que o voo batente e, portanto, necessita de menos massa muscular para sustentá-lo. Como um bônus adicional, a velocidades de cruzeiro normais, a asa é elevada (abduzida) na fase de subida por forças aerodinâmicas (Rayner 1988) - assim, contornando a necessidade da disposição do músculo supracoracoideu necessária para o voo batente de potência.
Ancestrais do Archaeopteryx
Além da ancestralidade terópode, foram sugeridos três outros grupos ancestrais para o Archae, os Ornithopoda, os Pseudosuchia e os Sphenosuchidae.
A teoria da ancestralidade ornitópoda baseava-se no tálus e no pélvis em vários dinossauros que eram supostamente próximos da condição encontrada nos pássaros. No entanto, a maioria dos táxons utilizados na teoria original de 1883-4 são agora conhecidos como sauriscianos e não ornitísquios. Embora houvesse uma tendência para a redução do hallux (dedão do pé) em alguns ornitísquios, ele nunca foi aparentemente opositor como é no Archae. Sugeriu-se que o Archae possuía um pélvis muito ornitísquio, no entanto, trabalhos recentes mostraram que isso é um artefato do movimento pós-mortem de elementos da região pélvica e há muito pouca semelhança com o pélvis ornitísquio. Com isso, o elo entre os ornitísquios e o Archae desaparece e, portanto, agora se pensa que não há nenhum elo entre eles.
Sugeriu-se que as aves surgiram de um grupo de répteis do Triássico Médio ou Tardio quase desconhecido, os Sphenosuchidae (Walker 1972, 1974), que também deram origem aos crocodilos. A teoria baseia-se em um único espécime de Sphenosuchus. No entanto, em suas configurações gerais, nem o crânio e as mandíbulas, nem a escápulo-coroide do Sphenosuchus sugerem remotamente afinidades com os Arqueossauros (Ostrom 1976). Mais recentemente, o principal defensor da hipótese crocodilo-ave afirmou que a hipótese "tornou-se tão tênue que é muito difícil sustentá-la." (Walker 1985, p. 133)
A ligação com os pseudossucianos foi muito favorecida no início deste século. No entanto, isso foi, em grande parte, baseado na reconstrução de Heilmann do espécime de Berlim, que desde então provou-se imprecisa. De acordo com Ostrom (1976 p. 159):
"Existem muito poucas semelhanças anatômicas entre Archaeopteryx e qualquer pseudossuciano. Na verdade, apenas em uma característica qualquer pseudossuciano se assemelha mais a Archaeopteryx do que qualquer terópode, sendo esta a razão entre a tíbia e o fêmur em Scleromochlus, Lagosuchus e Lagerpeton, onde a tíbia é de 20% a 30% mais longa que o fêmur. Entre os terópodes, apenas nos struthiomimídeos, Compsognathus, Microvenator e Deinonychus, a tíbia é mais longa que o fêmur, mas apenas em 10% a 15%. Em todas as outras características, a semelhança mais próxima com a morfologia preservada em Archaeopteryx é encontrada nos terópodes coelurosaurianos." [ênfase original]
Chiappe (1995) fornece uma excelente revisão da evolução inicial dos pássaros.
Protoavis
Algumas pessoas gostam de alegar que a descoberta de um fóssil de ave do Triássico do Texas (Protavis) prova que o Archaeopteryx não pode ser transicional entre dinossauros e aves porque o Protoavis antecede o Archaeopteryx em 75 milhões de anos. Isso, é claro, é uma tolice infundada, principalmente porque ninguém está alegando que o Archaeopteryx é a espécie transicional entre dinossauros e aves, mas apenas que o Archaeopteryx representa um grau de organização que a linhagem proposta passou para ir de dinossauros a aves. O Archaeopteryx, lamentavelmente, está numa rama, do ponto de vista evolutivo. Representa um ramo lateral, útil para fins comparativos, mas não está no centro das coisas. Então, mesmo que houvesse aves no Triássico, esse fato não diminuiria a importância do Archaeopteryx como indicador de que "sim, as aves poderiam ter evoluído de dinossauros".
No entanto, note o "se" na frase anterior. Existem problemas graves com o Protoavis. Na interpretação de Chatterjee (1991), Ostrom (1991) diz o seguinte [minha paráfrase]: O único material publicado do fóssil é um monográfico nas Philosophical Transactions da Royal Society de Londres. No entanto, este descreve apenas a cabeça. Esta está muito amassada e todas as peças foram extraídas da matriz, tornando a colocação precisa das peças questionável. A descrição é feita sob uma perspectiva aviar, sem contraponto (por exemplo, será que este é um dinossauro?) utilizado. O crânio está tão amassado que as características diagnósticas não foram preservadas. Portanto, o material publicado não suporta a visão de que este é um pássaro. De fato, uma visualização do fóssil por Ostrom (em circunstâncias admitidamente menos ideais) mostrou que as características diagnósticas que poderiam identificar o fóssil de qualquer forma estão muito amassadas e é duvidoso que qualquer afirmação definitiva possa ser sustentada pelo fóssil. Pode ser um pássaro, pode não ser.
Por favor, note que essa questionamento de Protoavis como um pássaro não é uma "reação malévola" evolucionista de "não pode ser um pássaro porque antecede Archae". Como foi apontado, mesmo que seja um pássaro, isso não diminui a importância evolutiva de Archae.
Conclusões
Archaeopteryx é um pássaro porque tinha penas. No entanto, manteve muitos caracteres dinossaúricos que não são encontrados em pássaros modernos, enquanto possuía certos caracteres encontrados em pássaros mas não em dinossauros. Por virtude deste facto, Archaeopteryx representa um exemplo de um grupo em transição - um representante que, embora à margem da transição de dinossauro para pássaro, um eco do evento real, ainda permite uma breve visão do possível mecanismo que levou à evolução dos pássaros e, pela sua própria existência, mostra que tal transição é possível.
Agradecimentos
Isso ocorreu como resultado de uma série de discussões com Rich Trott, que lutava contra o uso sistemático incorreto do Archae por certos opositores da evolução, e ainda assim conseguiu encontrar tempo para sugerir muitas melhorias neste post. As contribuições de Tom Holtz e Paul Davis também são reconhecidas. A maior parte da versão original foi convertida para o formato HTML para o arquivo talk.origins por Brett Vickers.
Esta é uma Contribuição Paleontológica da Universidade de Ediacara.
[Voltar às Archaeopteryx FAQs]
Referências
Abbott, A. (1992) Archaeopteryx fóssil desaparece de coleção particular. Nature, 357: 6.
Alexander, R.M. (1990) Animais. Cambridge University Press, Cambridge. 509pp.
Barsbold, R. et al. (1990) Oviraptorosauria. In The Dinosauria, Weishampel, Dodson & Osmolska (eds) pp 249-258.
Barthel, K.W.; Swinburne, N.H.M. & Conway Morris, S. (1990) Solnhofen. Um estudo em paleontologia mesozóica. Cambridge University Press, Cambridge. 236pp.
Bellairs, A.D'A. & Jenkin, C.R. (1960) O esqueleto dos pássaros. p. 241-300. In: Biologia e Fisiologia Comparada dos Pássaros, Vol. I, A.J. Marshal (ed.). Academic Press, Nova York.
Britt, B.B.; Makovicky, P.J.; Gauthier, J. & Bonde, N. (1998) Pneumatização pós-craniana em Archaeopteryx. Nature, 395: 374-376.
Brooks, B. (1993) Ossos pós-cranianos pneumáticos em dinossauros e outros arcosáurios. Tese de doutorado, Departamento de Geologia e Geofísica, Universidade de Calgary, Calgary, Alberta, Canadá.
Bryant, H.N. & Russell, A.P. (1993) A ocorrência de clavículas dentro dos Dinosauria: implicações para a homologia da furcula aviana e a utilidade da evidência negativa. Journal of Vertebrate Paleontology, 13(2):171-184.
Chatterjee, S. (1991) Anatomia craniana e relações de um novo pássaro do Triássico do Texas. Phil. Trans. R. Soc. Lond. B. 332: 277-342.
Chen Pei-Ji; Dong Zhi-Ming & Zhen Shuo-nan. (1998) Um dinossauro terópode excepcionalmente bem preservado da Formação Yixian, na China. Nature, 391: 147-152.
Chiappe, L.M. (1995) Os primeiros 85 milhões de anos da evolução das aves. Nature, 378: 349-355.
Chure, D.J. & Madsen, J.H. (1996) Sobre a presença de furculae em alguns terópodes não-maniraptóreos. Journal of Vertebrate Paleontology, 16: 573-577. Colbert, E. H. & Morales, M. (1991) Evolução dos vertebrados: uma história dos animais de espinha dorsal ao longo do tempo. 4ª ed. Wiley-Liss, Nova Iorque. 470 p.
de Beer, G. A. (1954) Archaeopteryx lithographica. British Museum of Natural History, London.
de Beer, G.A. (1956) Evolução dos Ratitas. Bulletin of the British Museum (Natural History): Zoologia, 4: 57-70.
Elzanowski, A. & Wellnhofer, P. (1996) Morfologia craniana de Archaeopteryx: Evidências do sétimo esqueleto. Journal of Vertebrate Paleontology 16 (1): 81-94
Gingerich, P.D. (1975) Significado evolutivo dos pássaros dentados do período mesozóico. Contribuições do Smithsonian à Paleobiologia, 27: 23-34.
Haubiltz, B.; Prokop, M.; Dohring, W.; Ostrom, J.H. & Wellnhofer, P. (1988) Tomografia computadorizada de Archaeopteryx. Paleobiology, 14: 206- 213.
Heilmann, G. (1926) A Origem dos Pássaros. Witherby, Londres. 208pp.
Jenkins, F.A. (1993) A evolução da articulação escapular da ave. American Journal of Science, 293A: 253-267.
Lambrecht, K. (1933) Handbuch der Palaeornithologie. Berlin. 1024 pp.
McFarland, W.N.; Pough, H.F.; Cade, H.F. & Heiser, J.B. (1985) Vertebrate Life, 2ª ed. Macmillan Publishing, Nova York. 636 pp.
McGowan, C. (1984) No Princípio: Um Cientista Mostra Por Que os Criacionistas Estão Errados. Prometheus, Buffalo. 208 pp.
Norberg, R.A. (1995) Assimetria das penas em Archaeopteryx. Nature 374: 221.
Orr, R.T. (1966) Vertebrate Biology, 2nd ed. W.B. Saunders, Philadelphia. 483pp.
Ostrom, J.H. (1970) Archaeopteryx: Aviso de um novo espécime. Science, 170: 537-538.
Ostrom, J.H. (1972) Descrição do espécime de Archaeopteryx no Museu Teyler, Haarlem. Proceedings Koninklijke Nederlandse Akademie Van Wetenschappen, B, 75: 289-305.
Ostrom, J.H. (1974) Archaeopteryx e a Origem do Voo. The Quarterly Review of Biology, 49(1)
Ostrom, J.H. (1976) Archaeopteryx e a origem dos pássaros. Biological Journal of the Linnean society, 8(2): 91-182.
Ostrom, J.H. (1980) Coelurus e Ornitholestes: São eles o mesmo? Em L. Jacobs (ed) Aspectos da História dos Vertebrados. 245-256.
Ostrom, J.H. (1991) Bird in the Bush. Nature, 353: 212
Ostrom, J.H. (1995) Biometria das asas e a origem do voo das aves. Neues Jahrbuch fur Geologie und Palaeontologie, 195: 253-266.
Padian, K. (1998) Quando um pássaro não é um pássaro? Nature, 393: 729-730.
Qiang Ji; Currie, P.J.; Norell, M.A. & Shu-an ji. (1998) Dois dinossauros emplumados da nordeste da China. Nature, 393: 753-761.
Rayner, J.M.V. (1988) A evolução do voo dos vertebrados. Biological Journal of the Linnean Society, 34: 269-287
Rietschel, S. (1985) In: The Beginnings of Birds, Hecht, M.K., Ostrom, J.H., Viohl, G. & Wellnhofer, P. (eds.). pp. 303-318. Freunde Des Jura-Museums, Eichstatt.
Romanoff, A.L. (1960) O Embrião Aves. Macmillan, Nova York. 1304pp.
Romer, A.S. (1950) Paleontologia Vertebrada. p 261. University of Chicago Press, Chicago.
Ruben, J. (1991) Fisiologia reptiliana e a capacidade de voo de Archaeopteryx. Evolution, 45: 1-17.
Speakman, J. R. (1993) Capacidades de voo em Archaeopteryx. Evolution, 47: 336-340.
Speakman, J.R. & Thomson, S.C. (1994) Capacidades de voo de Achaeopteryx. Nature, 370:514.
Tucker, B.W. (1938a) Morfologia evolutiva funcional: a origem das aves. p. 321-336. In: Evolução: Ensaios sobre Aspectos da Biologia Evolutiva, G.A. de Beer (ed.). Clarandon Press, Oxford.
Tucker, B.W. (1938b) Algumas observações sobre a teoria do Dr. Lowe sobre a relação dos Struthoines com os dinossauros e com outras aves. Proceedings of the 8th International Ornithological Congress, Oxford. 1934: 222-224.
Unwin, D M (1998) Penas, filamentos e dinossauros terópodes, Nature 391: 119-120.
Walker, A. D. (1972) Nova luz sobre as origens dos pássaros e crocodilos. Nature, 237: 257-263.
Walker, A. D. (1974) Evolução, orgânica. In Yearbook of Science and Technology, D.N. Lapedes (ed). pp. 177-179. McGraw-Hill, Nova York.
Walker, A. D. (1985) O crânio de Archaeopteryx. In The Beginnings of Birds, Hecht, M.K., Ostrom, J.H., Viohl, G. & Wellnhofer, P. (eds.). pp. 123-134. Freunde Des Jura-Museums, Eichstatt.
Wellnhofer, P. (1974) Das funfte skelettexemplar de Archaeopteryx. Palaeontographica A, 147: 169-216.
Wellnhofer, P. (1993) O sétimo exemplar de Archaeopteryx da calcária de Solnhofen. Archaeopteryx, 11: 1-47.
Whetstone, K. N. (1983) Caixa craniana de aves do Mesozoico: I. Nova preparação do "London" Archaeopteryx. Journal of Vertebrate Paleontology, 2: 439-452.
Witmer, L. M. (1990) O sistema de bolsas de ar craniofacial de aves do período Mesozoico (Aves). Zoological Journal of the Linnean Society, 100: 327-378.
[Voltar às Archaeopteryx FAQs]