|
por Raymond Sutera Copyright © 2001 [publicado em: 10 de agosto de 2001] Este artigo apareceu originalmente em Reports of the National Center for Science Education, uma publicação do The National Center for Science Education. |
 |
|
Como convencer um criacionista de que um fóssil é um fóssil transicional? Desistir? É uma questão-pegadinha. Você não consegue fazê-lo. Não é possível convencer alguém que já formou opinião. Mas, às vezes, é ainda pior. Às vezes, quando você aponta para um fóssil que cai no meio de uma lacuna e é um intermediário morfológico e cronológico excelente, recebe a resposta: “Bem, agora você tem duas lacunas em vez de uma! Você está recuando!”
Um dos desafios favoritos de antievolucionistas à existência de fósseis de transição é a suposta falta de formas de transição na evolução das baleias. Duane Gish, do Institute for Creation Research (ICR), costuma apresentar a transição “de bossie para sopro” para ridicularizar a ideia de que baleias poderiam ter evoluído de ancestrais terrestres de casco. Não há simplesmente formas de transição no registro fóssil entre os mamíferos marinhos e seus supostos ancestrais mamíferos terrestres... É bastante divertido, começando com vacas, porcos ou búfalos, tentar visualizar como os intermediários poderiam ter parecido. Começando com uma vaca, pode-se imaginar até uma linha de descendência que se extinguiu prematuramente, por algo que poderíamos chamar de falha de glândula mamária (Gish 1985: 78-9). Claro que, por muitos anos, o registro fóssil das baleias era bastante irregular, mas agora há numerosas formas de transição que ilustram a trajetória da evolução das baleias. Descobertas recentes de fósseis de baleias fornecem a evidência que convencerá um cético honesto. Contudo, a biologia evolutiva prevê não apenas a existência de ancestrais fósseis com certas características — ela também prevê que todas as outras disciplinas biológicas devem revelar também padrões de semelhança entre baleias, seus ancestrais e outros mamíferos correlacionados à evolução entre os grupos. Não deve surpreender que seja exatamente o que encontramos, e, como os achados em uma disciplina biológica, por exemplo bioquímica, são derivados sem referência aos achados em outra, por exemplo anatomia comparada, os cientistas consideram esses campos diferentes como evidências independentes da evolução das baleias. Como esperado, essas linhas independentes de evidência confirmam o padrão de evolução das baleias que anteciparíamos no registro fóssil. Para ilustrar essa abordagem, apresentarei evidências de múltiplas áreas para a origem das baleias a partir de mamíferos terrestres. Este texto examinará evidências mutuamente reforçadoras de nove áreas independentes de pesquisa. Claro, como ponto de partida, precisamos descrever o que faz uma baleia ser uma baleia. O que é uma baleia? Uma baleia é, antes de tudo, um mamífero — um vertebrado de sangue quente que usa seu metabolismo elevado para gerar calor e regular a temperatura interna. Fêmeas baleias têm filhotes vivos, que nutrem por glândulas mamárias. Embora baleias adultas não tenham cobertura de pelos corporais, elas adquirem pelos temporariamente como fetos, e algumas baleias adultas têm cerdas sensoriais ao redor da boca. Essas características são inequivocamente mamíferas. Mas uma baleia é um mamífero altamente especializado com muitos caracteres únicos que não são compartilhados com outros mamíferos — muitos deles nem sequer são compartilhados com outros mamíferos marinhos, como sirênios (peixes-boi e dugongos) e pinípedes (focas, leões-marinhos e morsas). Por exemplo, baleias têm corpos hidrodinâmicos que são espessos e arredondados, diferente dos corpos geralmente longos e esguios dos peixes. A cauda de uma baleia tem flúcidos horizontais, que são seu único meio de propulsão pela água. A nadadeira dorsal é endurecida por tecido conjuntivo, mas é carnosa e totalmente sem ossos de sustentação. As vértebras cervicais da baleia são encurtadas e pelo menos parcialmente fundidas em uma única massa óssea. As vértebras atrás do pescoço são numerosas e muito semelhantes umas às outras; os processos ósseos que conectam as vértebras são muito reduzidos, permitindo que as costas sejam muito flexíveis e produzam fortes impulsos das flidades de cauda. As barbatanas que permitem a direção da baleia são compostas de ossos de braço achatados e encurtados, ossos de pulso em forma de disco achatado e dedos alongados múltiplos. A articulação do cotovelo é praticamente imóvel, tornando a nadadeira rígida. Na cintura escapular, a escápula é achatada e não há clavícula. Algumas espécies de baleias ainda possuem uma pelve vestigial, e algumas têm membros posteriores muito reduzidos e não funcionais. A caixa torácica é muito móvel — em algumas espécies, as costelas estão totalmente separadas da coluna vertebral — o que permite que o tórax se expanda muito quando a baleia está respirando e permite que a caixa torácica se comprima em profundidade quando a baleia está mergulhando profundamente. O crânio também possui um conjunto de características únicas entre mamíferos. As mandíbulas se projetam para a frente, dando às baleias sua cabeça caracteristicamente longa, e os dois ossos mais anteriores da mandíbula superior (o maxilar e o pré-maxilar) são “telescópicos” para trás, às vezes cobrindo totalmente o topo do crânio. A migração posterior desses ossos é o processo pelo qual as aberturas nasais se moveram para o topo do crânio, criando sopros e deslocando o cérebro e o aparelho auditivo para a parte posterior do crânio. Os odontocetos (baleias dentadas) têm um único sopro, enquanto os misticetos (baleias de barbatanas) têm pares de sopros. Nos odontocetos, há uma assimetria pronunciada nos ossos telescópicos e no sopro que oferece um critério natural de classificação. Embora os dentes ocorram frequentemente em misticetos fetais, apenas odontocetos apresentam dentes em adultos. Esses dentes são sempre cones ou cavilhas simples; não são diferenciados por região ou função como os dentes de outros mamíferos. (As baleias não conseguem mastigar a comida; ela é moída no estômago anterior, ou papo muscular, que contém pedras.) Ao contrário do restante dos mamíferos, as baleias não têm glândulas lacrimais, glândulas da pele e senso olfativo. A audição é aguda, mas a orelha não possui abertura externa. A audição ocorre por vibrações transmitidas para um osso pesado e em forma de concha formado pela fusão de ossos do crânio (o periótico e os bullae auditivos). Essas, então, são as principais características das baleias. Algumas mostram claramente as adaptações distintas impostas às baleias por sua dedicação à vida marinha; outras conectam claramente as baleias a seus ancestrais terrestres. Outras mostram traços de descendência de um ancestral terrestre em comum com várias espécies antigas e modernas. A partir de todas essas características em conjunto, podemos reconstruir o caminho que a evolução das baleias percorreu de um ancestral terrestre até uma baleia moderna confinada a oceanos profundos. Pensando na ancestralidade da baleia Em 1693, John Ray registrou sua conclusão de que as baleias são mamíferos com base na similaridade das baleias com mamíferos terrestres (Barnes 1984). A discussão científica pré-darwinista girava em torno de se as baleias descendiam de mamíferos terrestres ou eram ancestrais deles. Darwin (1859) sugeriu que as baleias surgiram de ursos, esboçando um cenário no qual pressões seletivas poderiam fazer ursos evoluírem em baleias; envergonhado com críticas, ele removeu seus ursos nadadores hipotéticos de edições posteriores de Origin (Gould 1995). Mais tarde, Flower (1883) reconheceu que as baleias possuem características persistentes rudimentares e vestigiais características de mamíferos terrestres, confirmando assim que a direção da descendência era de espécies terrestres para marinhas. Com base na morfologia, Flower também vinculou as baleias aos ungulados; ele parece ter sido a primeira pessoa a fazer isso. No início do século XX, Eberhard Fraas e Charles Andrews sugeriram que creodontes (carnívoros primitivos, hoje extintos) eram os ancestrais das baleias (Barnes 1984). Mais tarde, WD Matthew, do American Museum of Natural History, postulou que as baleias descenderam de insetívoros, mas sua ideia nunca ganhou muito apoio (Barnes 1984). Ainda mais tarde, Everhard Johannes Slijper tentou combinar as duas ideias, alegando que as baleias descendiam do que Barnes chamou com precisão de “creodontes-cum-insetívoros”. No entanto, nenhum animal desse tipo foi encontrado. Mais recentemente, Van Valen (1966) e Szalay (1969) associaram as primeiras baleias aos condilarthes mesoníquios (um grupo agora extinto de ungulados carnívoros primitivos, nenhum maior que um lobo), com base em caracteres dentários. Evidências mais recentes confirmam essa avaliação. Assim Flower estava basicamente correto. A evidência A evidência de que as baleias descenderam de mamíferos terrestres é aqui dividida em nove partes independentes: paleontológica, morfológica, biologia molecular, vestigial, embrionária, geoquímica, paleoambiental, paleobiogeográfica e cronológica. Embora meu resumo da evidência não seja exaustivo, ele mostra que a visão atual da evolução das baleias é apoiada por pesquisa científica em várias disciplinas distintas. 1. Evidência paleontológica A evidência paleontológica vem do estudo da sequência fóssil de mamíferos terrestres até formas cada vez mais semelhantes a baleias até a aparição das baleias modernas. Embora as baleias iniciais (Arqueocetos) apresentem maior diversidade do que eu tenho espaço para discutir aqui, os exemplos nesta seção representam as tendências que vemos nesse táxon. Embora existam dois subordens modernos de baleias (odontocetos e misticetos), esta discussão se concentrará na origem das baleias como uma ordem de mamíferos e deixará de lado questões relacionadas à diversificação em subordens. Sinonyx Começamos com Sinonyx, um mesoníquio do tamanho de um lobo (um ungulado primitivo da ordem Condylarthra, que deu origem a artiodáctilos, perissodáctilos, proboscídeos e assim por diante) do final do Paleoceno, há cerca de 60 milhões de anos. Os caracteres que ligam Sinonyx às baleias, indicando assim que são aparentados, incluem um focinho alongado, um forame jugular aumentado e um básicoranium curto (Zhou e outros 1995). A contagem dentária era o número primitivo mamífero (44); os dentes eram diferenciados como os dentes heterodontos dos mamíferos atuais. Os molares eram dentes de corte muito estreitos, especialmente na mandíbula inferior, mas possuíam múltiplos cúspides. O alongamento do focinho está frequentemente associado à caça de peixes — todos os cetáceos caçadores de peixes, assim como os golfinhos, têm focinhos alongados. Essas características eram atípicas em mesoníquios, indicando que Sinonyx já estava desenvolvendo as adaptações que depois se tornariam a base do modo de vida especializado das baleias. Pakicetus O próximo fóssil da sequência, Pakicetus, é o mais antigo cetáceo e o primeiro arqueoceto conhecido. Ele é do início do Eoceno do Paquistão, há cerca de 52 milhões de anos (Gingerich e outros 1983). Embora seja conhecido apenas por restos cranianos fragmentários, esses restos são muito diagnósticos e são definitivamente intermediários entre Sinonyx e baleias posteriores. Isso é especialmente o caso dos dentes. Os molares superiores e inferiores, com múltiplos cúspides, ainda se assemelham aos de Sinonyx, mas os pré-molares tornaram-se dentes triangulares simples compostos de uma única cúspide serrilhada nas bordas frontal e posterior. Os dentes de baleias posteriores mostram ainda mais simplificação em triângulos serrilhados simples, como os de tubarões carnívoros, indicando que os dentes de Pakicetus eram adaptados à caça de peixes.  Gingrich and others (1983) published this reconstruction of the skull of Pakicetus inachus (redrawn for RNCSE by Janet Dreyer). Um crânio bem preservado mostra que Pakicetus era definitivamente um cetáceo com uma cápsula craniana estreita, crista sagital alta e estreita e cristas lambdoides proeminentes. Gingerich e outros (1983) reconstruíram um crânio composto com cerca de 35 centímetros de comprimento. Pakicetus não ouvia bem debaixo d’água. Seu crânio não tinha nem bullae timpânicas densas nem seios que isolassem a área auditiva esquerda da direita — uma adaptação de baleias posteriores que permite audição direcional debaixo d’água e impede a transmissão de sons pelo crânio (Gingerich e outros 1983). Todas as baleias vivas têm seios preenchidos com espuma junto com bullae timpânicas densas que criam um contraste de impedância para que possam separar sons vindos de diferentes direções. Não há também evidência em Pakicetus de vascularização do ouvido médio, necessária para regular a pressão no ouvido médio durante o mergulho (Gingerich e outros 1983). Portanto, Pakicetus provavelmente era incapaz de atingir mergulhos de qualquer profundidade significativa. Essa avaliação paleontológica do nicho ecológico de Pakicetus é totalmente consistente com a evidência geoquímica e paleoambiental. Quando se trata de audição, Pakicetus era mais terrestre do que aquático, mas a forma de seu crânio era definitivamente de cetáceo, e seus dentes estavam entre os estados ancestral e moderno.  Zhou and others (1995) published this reconstruction of the skull of Sinonyx jiashanensis (redrawn for RNCSE by Janet Dreyer). Ambulocetus Na mesma região onde Pakicetus foi encontrado, mas em sedimentos cerca de 120 metros mais acima, Thewissen e colegas (1994) descobriram Ambulocetus natans, “a baleia caminhante que nada”, em 1992. Datado do início ao meio do Eoceno, há cerca de 50 milhões de anos, Ambulocetus é um fóssil verdadeiramente notável. Era claramente um cetáceo, mas também tinha pernas funcionais e um esqueleto que ainda permitia algum grau de locomoção terrestre. A conclusão de que Ambulocetus podia caminhar usando os membros posteriores é apoiada por possuir um fêmur grande e robusto. No entanto, como o fêmur não tinha os pontos de fixação grandes o suficiente para os músculos da locomoção, não poderia ser um caminhante muito eficiente. Provavelmente só podia andar de modo que os leões-marinhos modernos conseguem andar — girando os pés traseiros para a frente e cambaleando no solo com a ajuda das patas dianteiras e da flexão da coluna. Ao andar, suas enormes patas dianteiras devem ter apontado para o lado em alguma medida, pois, se apontassem para a frente, interfeririam entre si. Os membros anteriores também eram intermediários tanto na estrutura quanto na função. A ulna e o rádio eram fortes e capazes de suportar o peso do animal em terra. O cotovelo forte era robusto, mas estava inclinado para trás, tornando possíveis impulsos para trás do antebraço para a natação. No entanto, os punhos, ao contrário dos de baleias modernas, eram flexíveis. É óbvio pela anatomia da coluna vertebral que Ambulocetus devia nadar com a espinha oscilando para cima e para baixo, impulsionado pelas patas traseiras, orientadas para trás. Como em outros mamíferos aquáticos que usam esse método de natação, as patas traseiras eram bastante grandes. De modo incomum, os dedos das patas traseiras terminavam em casco, exibindo assim a ascendência ungulada do animal. A única vértebra caudal encontrada é longa, tornando provável que a cauda também fosse longa. As vértebras cervicais eram relativamente longas, em comparação com as de baleias modernas; Ambulocetus devia ter um pescoço flexível. O crânio de Ambulocetus era bastante cetáceo (Novacek 1994). Tinha um focinho longo, dentes muito semelhantes aos de arqueocetos posteriores, um arco zigomático reduzido e uma bula timpânica (que sustenta o tímpano) que era mal fixada ao crânio. Embora Ambulocetus aparentemente não tivesse sopro, os demais recursos cranianos qualificam Ambulocetus como cetáceo. As características pós-craniais são claramente de adaptação de transição para o ambiente aquático. Assim, Ambulocetus é melhor descrito como um anfíbio do tamanho de um leão-marinho, piscívoro, que ainda não estava totalmente desconectado da vida terrestre de seus ancestrais. Rodhocetus No Eoceno médio (46–47 milhões de anos atrás), Rodhocetus levou todas essas mudanças ainda mais adiante, mantendo ainda vários traços terrestres primitivos (Gingerich e outros 1994). É o arqueoceto mais antigo de que todas as vértebras torácicas, lombares e lombossacras foram preservadas. As vértebras lombares tinham espinhos neurais mais altos do que em baleias anteriores. O tamanho dessas projeções no topo das vértebras onde músculos se fixam indica que Rodhocetus desenvolvera uma cauda poderosa para natação. 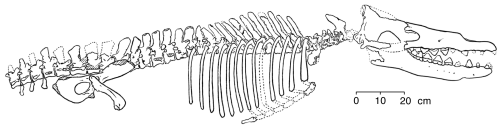 Gingrich and others (1994) published this reconstruction of the skeleton of Rodhocetus kasrani (redrawn for RNCSE by Janet Dreyer). Em outra parte da coluna vertebral, as quatro grandes vértebras sacrais estavam não fundidas. Isso dava à coluna mais flexibilidade e permitia uma propulsão mais potente durante a natação. Também é provável que Rodhocetus tivesse uma aleta caudal, embora essa característica não esteja preservada nos fósseis conhecidos: possuía características — vértebras cervicais encurtadas, vértebras caudais proximais pesadas e robustas e grandes espinhos dorsais nas vértebras lombares para grandes inserções de músculos axiais e caudais — que, nas baleias modernas, estão associadas ao desenvolvimento e uso de barbatanas caudais. Em resumo, Rodhocetus deve ter sido um excelente nadador de cauda, e é o fóssil de baleia mais antigo comprometido com esse tipo de natação. A pelve de Rodhocetus era menor que a de seus predecessores, mas ainda estava conectada às vértebras sacrais, o que significa que Rodhocetus ainda podia andar em terra em alguma medida. No entanto, a asa do osso ilíaco era curta em comparação com a dos mesoníquios, o que gerava um impulso muscular menos poderoso do quadril durante a caminhada, e o fêmur era cerca de 1/3 mais curto que os de Ambulocetus, de modo que Rodhocetus provavelmente não se movia tão bem em terra quanto seus predecessores (Gingerich e outros 1994). O crânio de Rodhocetus era relativamente grande em comparação com o restante do esqueleto. Os pré-maxilares e as mandíbulas inferiores se estendiam para frente ainda mais que seus predecessores, alongando o crânio e tornando-o ainda mais cetáceo. Os molares têm coroas mais altas que em baleias anteriores e estão altamente simplificados. Os molares inferiores são mais altos do que largos. Há uma diferenciação reduzida entre os dentes. Pela primeira vez, as narinas se moveram para trás ao longo do focinho e ficam acima dos dentes caninos, mostrando a evolução do sopro. As bullae auditivas são grandes e feitas de osso denso (características únicas de cetáceos), mas aparentemente não continham os seios típicos das baleias mais recentes, o que põe em dúvida se Rodhocetus possuía audição direcional debaixo d’água. No geral, Rodhocetus mostrou melhorias em relação às baleias anteriores por meio de tórax profundo e esguio, cabeça mais longa, maior flexibilidade vertebral e musculatura caudal expandida. O aumento da flexibilidade e da força nas costas e na cauda, com a diminuição correspondente da força e do tamanho dos membros, indicou que era um bom nadador de cauda com capacidade reduzida de andar em terra. Basilosaurus O fóssil de baleia especialmente conhecido Basilosaurus representa o próximo grau evolutivo na evolução das baleias (Gingerich 1994). Viveu no final do Eoceno e na parte mais recente do Eoceno médio (35-45 milhões de anos atrás). Basilosaurus era um animal longo, fino e serpentiforme que originalmente se pensava ser de um monstro marinho (daí seu nome, que significa “lagarto rei”). Seu comprimento corporal extremo (cerca de 15 metros) parece dever-se a uma característica única entre as baleias; suas 67 vértebras são tão longas em comparação com outras baleias da época e com baleias modernas que provavelmente representam uma especialização que o distingue da linhagem que deu origem às baleias modernas. O que torna Basilosaurus uma baleia particularmente interessante, entretanto, é a anatomia distintiva de seus membros posteriores (Gingerich e outros 1990). Possuía uma cintura pélvica quase completa e um conjunto de ossos de membros posteriores. Os membros eram pequenos demais para propulsão eficaz, com menos de 60 cm nesse animal de 15 metros, e a cintura pélvica estava completamente isolada da espinha, tornando impossível o suporte de peso. Reconstruções do animal colocaram suas patas fora do corpo — uma configuração que representaria uma forma intermediária importante na evolução das baleias. Embora uma barbatana caudal nunca tenha sido encontrada (já que barbatanas caudais não têm ossos e é improvável fossilizarem), Gingerich e outros (1990) observaram que a coluna vertebral de Basilosaurus compartilha características de baleias que têm barbatanas caudais. As vértebras caudais e cervicais são mais curtas do que as regiões torácicas e lombares, e Gingerich e outros (1990) tomam essas proporções vertebrais como evidência de que Basilosaurus provavelmente também tinha barbatana caudal. Mais evidências de que Basilosaurus passava a maior parte do tempo na água vêm de outra mudança importante no crânio. Este animal tinha uma grande narina única que se migrara uma curta distância para trás até um ponto correspondente ao terço posterior da série dentária. O deslocamento da ponta do focinho para uma posição mais próxima ao topo da cabeça é característico apenas de mamíferos que vivem em ambientes marinhos ou aquáticos. Dorudon Dorudon foi contemporâneo de Basilosaurus no final do Eoceno (cerca de 40 milhões de anos atrás) e provavelmente representa o grupo mais provável de ancestral das baleias modernas (Gingerich 1994). Dorudon não tinha as vértebras alongadas de Basilosaurus e era muito menor (cerca de 4-5 metros de comprimento). A dentição de Dorudon era semelhante à de Basilosaurus; seu crânio, em comparação com os crânios de Basilosaurus e das baleias anteriores, era um pouco abobadado (Kellogg 1936). Dorudon também ainda não tinha a anatomia craniana que indica a presença do aparato necessário para ecolocalização (Barnes 1984).  Gingrich and Uhen (1996) published this reconstruction of the skeleton of Dorudon atrox (redrawn for RNCSE by Janet Dreyer). Basilosaurus e Dorudon eram baleias totalmente aquáticas (como Basilosaurus, Dorudon tinha membros posteriores muito pequenos que podem ter se projetado um pouco além da parede corporal). Já não estavam ligados à terra; de fato, não teriam conseguido se mover em terra. Seu tamanho e a ausência de membros que suportassem seu peso os tornavam mamíferos aquáticos obrigatórios, uma tendência que é elaborada e reforçada por táxons subsequentes. Claramente, mesmo se olharmos apenas para a evidência paleontológica, a alegação criacionista de “Sem intermediários fósseis!” está errada. Na verdade, no caso das baleias, temos vários, belamente dispostos em ordem morfológica e cronológica. Ao resumir a evidência paleontológica, observamos mudanças consistentes que indicam uma série de adaptações de ambientes mais terrestres para mais aquáticos conforme avançamos da espécie mais ancestral para a mais recente. Essas mudanças afetam a forma do crânio, a forma dos dentes, a posição das narinas, o tamanho e a estrutura dos membros anteriores e posteriores, o tamanho e a forma da cauda e a estrutura do ouvido médio em relação à audição direcional debaixo d’água e ao mergulho. A evidência paleontológica registra uma história de adaptação crescente à vida na água — não apenas a qualquer modo de vida aquático, mas ao modo de vida vivido pelas baleias contemporâneas. 2. Evidência morfológica A análise das características morfológicas compartilhadas por fósseis de baleias e ungulados vivos torna ainda mais clara sua ascendência comum. Por exemplo, a anatomia do pé de Basilosaurus alinha as baleias com artiodáctilos (Gingerich e outros 1990). O eixo de simetria do pé nesses fósseis de baleias fica entre os 3º e 4º dígitos. Essa disposição é chamada paraxônica e é característica dos artiodáctilos, baleias e condilarthes, e raramente encontrada em outros grupos (Wyss 1990). Outro exemplo envolve o incus (a “bigorna” do ouvido médio). O incus de Pakicetus, preservado em ao menos um espécime, é morfologicamente intermediário em todos os caracteres entre o incus das baleias modernas e o de artiodáctilos modernos (Thewissen e Hussain 1993). Além disso, a articulação entre o malleus (martelo) e o incus da maioria dos mamíferos está orientada em ângulo entre o meio e a parte frontal do animal (rostromedialmente), enquanto em baleias modernas e em ungulados, está orientada em ângulo entre o lado e a frente (rostrolateralmente). Em Pakicetus, o primeiro fóssil cetáceo, a articulação está orientada rostralmente (intermediária em posição entre as condições ancestral e derivada). Assim, a articulação claramente girou em direção ao meio a partir da condição ancestral em mamíferos terrestres (Thewissen e Hussain 1993); Pakicetus nos oferece um retrato da transição. 3. Evidência biológica molecular A hipótese de que baleias descendem de mamíferos terrestres prevê que baleias atuais e mamíferos terrestres vivos intimamente relacionados devem mostrar semelhanças em sua biologia molecular aproximadamente em proporção à atualidade de seu ancestral comum. Ou seja, as baleias deveriam ser mais parecidas, em biologia molecular, com grupos de animais com os quais compartilham um ancestral comum mais recente do que com outros animais que exibem semelhanças convergentes em morfologia, ecologia ou comportamento. Em contraste, o criacionismo não tem base científica para prever quais padrões de semelhança deveriam ocorrer, pois não há modo científico de prever como o criador decidiu distribuir semelhanças moleculares entre espécies. Estudos moleculares de Goodman e outros (1985) mostram que as baleias estão mais estreitamente relacionadas aos ungulados do que a todos os outros mamíferos — resultado compatível com as expectativas evolutivas. Esses estudos examinaram mioglobina, cristalina alpha-lens e citocromo c em 46 espécies diferentes de mamíferos. Miyamoto e Goodman (1986) ampliaram posteriormente o número de sequências proteicas incluindo alpha e beta hemoglobinas e ribonuclease; também aumentaram o número de mamíferos incluídos no estudo para 72. Os resultados foram os mesmos: as baleias estão claramente incluídas entre os ungulados. Outros estudos moleculares de diversos genes, proteínas e enzimas por Irwin e outros (1991), Irwin e Arnason (1994), Milinkovitch (1992), Graur e Higgins (1994), Gatesy e outros (1996) e Shimamura e outros (1997) também identificaram baleias como próximas dos artiodáctilos, embora haja diferenças nos detalhes entre os estudos. Ao colocar as baleias próximas e até firmemente dentro dos Artiodactyla, esses estudos moleculares confirmam as previsões feitas pela teoria evolutiva. Esse padrão de semelhanças bioquímicas precisa estar presente se baleias e ungulados, especialmente os artiodáctilos, compartilham um ancestral comum próximo. O fato de essas semelhanças estarem presentes é, portanto, evidência forte para a descendência comum de baleias e ungulados. 4. Evidência vestigial As características vestigiais das baleias nos dizem duas coisas. Elas nos mostram que as baleias, como tantos outros organismos, possuem características que não fazem sentido sob uma perspectiva de design — não têm função atual, exigem energia para serem produzidas e mantidas e podem ser prejudiciais ao organismo. Elas também nos mostram que as baleias carregam consigo um pedaço de seu passado evolutivo, destacando uma história de ascendência terrestre. Baleias modernas frequentemente retêm vestígios em forma de bastão de ossos pélvicos, fêmures e tíbias, todos embutidos na musculatura de suas paredes corporais. Esses ossos são mais pronunciados em espécies anteriores e menos pronunciados em espécies posteriores. Como exemplo de Basilosaurus mostra, baleias de idade intermediária têm pélvices e ossos dos membros posteriores de tamanho intermediário. As baleias também retêm um número de estruturas vestigiais em seus órgãos de sensibilidade. Baleias modernas têm apenas nervos olfativos vestigiais. Além disso, em baleias modernas, o meato auditivo (a abertura externa do canal auditivo) está fechado. Em muitas, tem apenas o tamanho de um fio fino, de cerca de 1 mm de diâmetro, e costuma ficar fechado no meio. Todas as baleias têm vários pequenos músculos dedicados a orelhas externas inexistentes, que aparentemente são resquícios de um tempo em que podiam mover as orelhas — um comportamento tipicamente usado por animais terrestres para audição direcional. O diafragma nas baleias é vestigial e tem muito pouco músculo. As baleias usam o movimento para fora das costelas para encher os pulmões com ar. Finalmente, Gould (1983) relatou várias ocorrências de cachalotes capturados com membros posteriores visíveis e salientes. Da mesma forma, golfinhos foram observados com pequenas nadadeiras pélvicas, embora provavelmente não fossem suportadas por ossos de membros como nesses raros cachalotes. E algumas baleias, como as belugas, possuem aurículas auriculares rudimentares — uma característica que não pode servir a qualquer propósito em um animal sem orelha externa e que pode reduzir a eficiência de natação ao aumentar o arrasto hidrodinâmico durante a natação. Embora essa lista esteja de modo algum esgotada, é inquestionável que as baleias têm uma riqueza de características vestigiais herdadas de seus ancestrais terrestres. 5. Evidência embrionária Como as características vestigiais, as características embrionárias também nos contam duas coisas. Primeiro, o embrião da baleia desenvolve um número de características que depois abandona antes de atingir sua forma final. Como o criacionismo pode explicar tal processo aparentemente sem sentido, construindo estruturas apenas para abandoná-las ou destruí-las depois? Darwin (1859) fez a mesma pergunta. Não faria mais sentido os embriões alcançarem rapidamente suas formas adultas diretamente? Parece irrazoável para um designer perfeito ou criador enviar o embrião por caminho tão tortuoso, mas a evolução requer que novas características sejam construídas sobre a base de características anteriores que depois ele modificaria ou descartaria. Segundo, a embriologia da baleia, examinada em detalhes, também oferece evidência de sua ancestralidade terrestre. Tanto os embriões quanto os animais adultos, as baleias são verdadeiros depósitos de restos, digamos, de antigas características descartadas que não têm mais uso para elas. Muitas baleias, ainda no útero, começam a desenvolver pelos corporais. No entanto, nenhuma baleia moderna retém pelos corporais após o nascimento, exceto algumas cerdas no focinho e pelos ao redor dos sopros usados como cerdas sensoriais em algumas espécies. O fato de as baleias possuírem os genes para produzir pelos corporais mostra que seus ancestrais tinham pelos corporais. Em outras palavras, seus ancestrais eram mamíferos ordinários. Em muitos embriões de baleia, brotos dos membros posteriores externos são visíveis por um período, mas desaparecem quando a baleia cresce. Também são visíveis no embrião aurículos auriculares rudimentares, que desaparecem antes do nascimento (exceto naqueles que os carregam como atavismos raros). E, em algumas baleias, os lóbulos olfativos do cérebro existem apenas no feto. O embrião da baleia começa com as narinas no lugar usual dos mamíferos, no ápice do focinho. Mas durante o desenvolvimento, as narinas migram para seu local final no topo da cabeça para formar o sopro (ou sopros). Também podemos compreender a evolução dentro das baleias por meio de sua embriologia. Sabemos que as baleias de barbatanas (misticetos) evoluíram das baleias dentadas: alguns embriões de baleias de barbatanas começam a desenvolver dentes. Como com os pelos corporais, os dentes desaparecem antes do nascimento. Como não há uso para dentes no útero, apenas a herança de um ancestral comum faz sentido; não há razão para o designer inteligente ou criador especial fornecer dentes aos embriões de baleias. Assim, temos mais uma área independente em total concordância com a tese geral — de que as baleias possuem características que as conectam com ancestrais mamíferos terrestres, em particular os mamíferos de casco. 6. Evidência geoquímica As baleias mais antigas viveram em habitats de água doce, mas os ancestrais das baleias modernas moveram-se para habitats de água salgada e, portanto, tiveram de se adaptar a beber água salgada. Como água doce e água salgada têm razões isotópicas de oxigênio um pouco diferentes, podemos prever que a transição será registrada nos restos esqueléticos das baleias — os mais duráveis entre os quais estão os dentes. Certamente, dentes fósseis das baleias mais antigas têm proporções menores de oxigênio pesado para oxigênio leve, indicando que os animais bebiam água doce (Thewissen e outros 1996). Dentes fósseis posteriores de baleias têm proporções maiores de oxigênio pesado para oxigênio leve, indicando que beberam água salgada. Isso reforça absolutamente a inferência retirada de toda a outra evidência discutida aqui: os ancestrais das baleias modernas se adaptaram de habitats terrestres para habitats de água salgada por intermédio de habitats de água doce. 7. Evidência paleoambiental A evolução faz outras previsões sobre a história dos táxons com base na visão “macro” dos fósseis em um contexto ambiental mais amplo. A sequência de fósseis de baleias e suas mudanças também deve se relacionar às mudanças observadas nos registros fósseis de outros organismos no mesmo tempo e em ambientes semelhantes. Os fósseis de outros organismos associados aos fósseis de baleias indicam o ambiente em que as baleias viveram. Além disso, essa evidência deve ser consistente com a evidência das outras áreas de estudo. Espera-se encontrar evidência de uma série de ambientes de transição, de totalmente terrestre para totalmente marinho, ocupada pela série de espécies de baleias no registro fóssil. A morfologia de Sinonyx indica que era totalmente terrestre. Portanto, não deveria surpreender que seus fósseis sejam encontrados associados a fósseis de outros animais terrestres. Pakicetus provavelmente passava muito tempo na água em busca de alimento. Embora a fauna mamífera encontrada com Pakicetus consistisse de roedores, morcegos, vários artiodáctilos, perissodáctilos e proboscídeos, e até um primata (Gingerich e outros 1983), também há animais aquáticos como caracóis, peixes, tartarugas e crocodilianos. Além disso, o sedimento associado a Pakicetus mostra evidência de fluxo ou corrente, geralmente associado a solos que são transportados pela água. A evidência paleoambiental mostra claramente que Pakicetus viveu em ambiente terrestre úmido de baixa altitude, fazendo incursões ocasionais em água doce. Interessantemente, dentes decíduos e permanentes do animal são encontrados nesses sedimentos com frequência semelhante, apoiando a ideia de que Pakicetus dava à luz em terra. Os sedimentos em que Ambulocetus foi encontrado contêm impressões de folhas além de fósseis do caramujo-turritela Turritella e outros moluscos marinhos. Claramente, a presença de tais fósseis deve significar que o fóssil de Ambulocetus foi encontrado onde antes havia um mar raso — embora folhas possam ser levadas ao mar e se fossilizarem lá, moluscos marinhos não seriam encontrados em terra. Rodhocetus é encontrado em folhelhos verdes depositados na zona nerítica profunda (equivalente à parte externa da plataforma continental). Como folhelhos verdes estão associados a águas de fundo com baixo teor de oxigênio, Rodhocetus deve ter vivido a profundidades maiores que qualquer cetáceo anterior. O fato de ser encontrado em associação com foraminíferos planctônicos e outros microfósseis concorda com essa determinação de profundidade. Basilosaurus e Dorudon foram encontrados em variedade de tipos de sedimento (Kellogg 1936), indicando que eram de ampla distribuição e capazes de viver em águas profundas e também rasas. A partir da evidência paleoambiental, podemos ver claramente que, conforme as baleias evoluíam, elas avançavam para águas mais profundas e tornavam-se progressivamente liberadas dos ambientes terrestres e de margem. 8. Evidência paleobiogeográfica A evidência geográfica também é consistente com os padrões de distribuição esperados para o primeiro aparecimento das baleias e posterior expansão geográfica. Esperaríamos que espécies terrestres tivessem distribuição geográfica mais restrita do que espécies marinhas, que têm essencialmente todo o oceano como faixa geográfica. A distribuição de Sinonyx está restrita à Ásia central. Espécimes de Pakicetus só foram encontrados no Paquistão; Ambulocetus e Rodhocetus parecem igualmente restritos. Em contraste, Basilosaurus e Dorudon, representando as baleias mais adaptadas a viver no mar aberto, são encontrados em uma área muito maior. Seus fósseis foram achados tão longe da Ásia meridional quanto Geórgia, Louisiana e British Columbia. Durante o Eoceno, a maioria das áreas em que fósseis de baleias posteriores foram encontrados estava relativamente próxima umas das outras. De fato, a maioria delas está ao longo da margem externa de um mar antigo chamado Tetis, cujos remanescentes hoje são o Mediterrâneo, o Cáspio, o Negro e o Mar de Aral. A distribuição biogeográfica de fósseis de baleias corresponde ao padrão previsto pela evolução: as baleias são inicialmente encontradas em uma área geográfica relativamente pequena e não se tornaram distribuídas pelo mundo até depois de evoluírem em animais totalmente aquáticos que já não estavam mais ligados à terra. 9. Evidência cronológica A última corrente de evidência em nosso quadro mutuamente consistente sobre as origens das baleias vem de uma consideração de por que as baleias se originaram quando o fizeram. A evolução é uma resposta a desafios e oportunidades ambientais. Durante o início do Cenozoico, os mamíferos foram apresentados a um novo conjunto de oportunidades de radiação e diversificação devido, em parte, ao vazio deixado pelas extinções em massa no fim do Período Cretáceo. Como os répteis não predominavam mais, surgiram novas formas pelas quais os mamíferos poderiam ganhar a vida. No caso específico das baleias, os répteis nadadores dos oceanos do mundo não conseguiam mais manter os mamíferos afastados da costa. Antes das extinções do fim do Cretáceo, os répteis marinhos mesozoicos, como pliossauros, ictiossauros, mosassauros e crocodilos marinhos, podem muito bem ter devorado qualquer mamífero que se afastasse da costa em busca de alimento. Uma vez que esses predadores desapareceram, a evolução produziu rapidamente mamíferos, incluindo baleias, tão à vontade nos mares como antes estavam na terra. A transição levou cerca de 10 a 15 milhões de anos para produzir baleias totalmente aquáticas de mergulho profundo com audição direcional subaquática. A evolução prevê que as baleias não poderiam ter aparecido e radido com sucesso antes do Eoceno e que os mamíferos deveriam ter radiado para ambientes marinhos assim como radiaram para uma ampla variedade de outros ambientes deixados pelos répteis no fim do Cretáceo. Conclusão Tomados em conjunto, todas essas evidências apontam para uma única conclusão — de que as baleias evoluíram de mamíferos terrestres. Vimos que há nove áreas de estudo independentes que fornecem evidências de que as baleias compartilham um ancestral comum com mamíferos de casco. O poder da evidência de áreas de estudo independentes que apoiam a mesma conclusão torna totalmente irrazoável a refutação por cenários de criação especial, incredulidade pessoal, o argumento da ignorância ou cenários de “design inteligente”. A única conclusão científica plausível é que as baleias realmente evoluíram de mamíferos terrestres. Portanto, não importa quanta gente antievolucionista reclame de quão impossível é para mamíferos peludos terrestres evoluírem em baleias totalmente aquáticas, a própria evidência as silencia. Este é o poder de usar linhas de evidência mutuamente reforçadoras e independentes. Espero que se torne uma arma importante para derrubar objeções antievolucionistas infundadas e apoiar o pensamento evolutivo no público em geral. Assim funciona a ciência real, e devemos enfatizar o processo de inferência científica conforme apontamos as conclusões que os cientistas extraem da evidência — de que as previsões concordantes de campos independentes de estudo científico confirmam o mesmo padrão de ancestralidade das baleias. Agradecimentos Gostaria de agradecer ao Dr. Philip Gingerich por sua assistência e revisão deste artigo. Referências Barnes LG. Search for the first whale. Oceans 1984 March-April; 17 (2): 20-3. Darwin CR. On the Origin of Species. New York: Random House, 1859. Flower WH. On the arrangement of the orders and families of existing Mammalia. Proceedings of the Zoological Society of London 1883 Aug; 1: 178-86. Gatesy J, Hayashi C, Cronin MA, Arctander P. Evidence from milk casein genes that cetaceans are close relatives of hippopotamid artiodactyls. Molecular Biology and Evolution 1996; 13 (7): 954-63. Gingerich P. The whales of Tethys. Natural History 1994 April; 103 (4): 86-8. Gingerich P, Raza SM, Arif M, Anwar M, Zhou X. New whale from the Eocene of Pakistan and the origin of cetacean swimming. Nature 1994; 368: 844-7. Gingerich P, Smith BH, Simons EL. Hind limbs of Eocene Basilosaurus: evidence of feet in whales. Science 1990; 249: 154-7. Gingerich P, Wells NA, Russell DE, Shah SMI. Origin of whales in epicontinental remnant seas: new evidence from the early Eocene of Pakistan, Science 1983; 220: 403-6. Gish DT. Evolution: The Challenge of the Fossil Record. El Cajon (CA): Creation-Life Publishers, 1985. Goodman M, Czelusniak J, Beeber J. Phylogeny of primates and other eutherian orders: a cladistic analysis using amino acid and nucleotide sequence data. Cladistics 1985; 1 (2): 171-85. Gould SJ. Hen's teeth and horse's toes. In: Hen's Teeth and Horse's Toes. Norton: New York, 1983. p 177-86. Gould SJ. Hooking leviathan by its past. In: Dinosaur in a Haystack, New York: Harmony Books, 1995. p 359-76. Graur D, Higgins DG. Molecular evidence for the inclusion of cetaceans within the order Artiodactyla. Molecular Biology and Evolution 1994; 11 (3): 357-64. Irwin DM, Arnason U. Cytochrome b gene of marine mammals: phylogeny and evolution. Journal of Mammalian Evolution 1994; 2 (1): 37-55. Irwin DM, Kochner TD, Wilson AC. Evolution of the cytochrome b gene of mammals. Journal of Molecular Evolution 1991; 32: 128-44. Kellogg R. A Review of the Archaeoceti. Washington DC: Carnegie Institute, 1936. Matthews LH. The Natural History of the Whale. New York: Columbia University Press, 1978. Milinkovitch MC. DNA-DNA hybridizations support ungulate ancestry of cetacea. Journal of Evolutionary Biology 1992; 5: 149-60 Milinkovitch MC, Orti G, Meyer A. Revised phylogeny of whales suggested by mitochondrial ribosomal DNA sequences. Nature 1993; 361: 346-8. Miyamoto MM, Goodman M. Biomolecular systematics of eutherian mammals: phylogenetic patterns and classification. Systematic Zoology 1986; 35 (2):230-40. Novacek M. Mammalian phylogeny: shaking the tree. Nature 1992; 356: 121-5. Novacek M. Whales leave the beach. Nature 1994; 368: 807. Shimamura M, Yasue H, Ohshima K, Abe H, Kato H, Kishiro T, Goto M, Munechika I, Okada N. Molecular evidence from retroposons that whales form a clade within even-toed ungulates. Nature 1997; 388: 666-9. Szalay FS. Origin and evolution of function of the mesonychid condylarth feeding mechanism. Evolution 1969; 23: 703-20. Thewissen JGM, Hussain ST. Origin of underwater hearing in whales. Nature 1993; 361: 444-5. Thewissen JGM, Hussain ST, Arif M. Fossil evidence for the origin of aquatic locomotion in Archaeocete whales. Science 1994; 263: 210-2. Thewissen JGM, Roe LJ, O'Neill JR, Hussain ST, Sahni A, Bajpal S. Evolution of cetacean osmoregulation. Nature 1996; 381: 379-80. Van Valen L. Deltatheridia, a new order of mammals. Bulletin of the American Museum of Natural History 1966; 132: 1-126. Wyss A. Clues to the origin of whales. Nature 1990; 347: 428-9. Zhou X, Zhai R, Gingerich P, Chen L. Skull of a new Mesonychid (Mammalia, Mesonychia) from the late Paleocene of China. Journal of Vertebrate Paleontology 1995; 15 (2): 387-400. Endereço do autor: Ray Sutera 81 Franklin Ave. Ocean Grove NJ 07756-1128 rsutera@aol.com |