Ist das Komplementsystem irreduzibel komplex?
von Mike Coon![]()
"Unter irreduzibler Komplexität verstehe ich ein einzelnes System, das aus mehreren interagierenden Teilen besteht, die zur Grundfunktion beitragen, und bei dem das Entfernen eines beliebigen Teils dazu führt, dass das System effektiv seine Funktion einstellt. Ein irreduzibel komplexes System kann nicht schrittweise durch geringfügige, aufeinanderfolgende Modifikationen eines Vorläufersystems erzeugt werden, da jeder Vorläufer eines irreduzibel komplexen Systems per Definition nicht funktionsfähig ist."
-- Michael Behe
Dieser Aufsatz soll einige Elemente der hervorragenden FAQ von Keith Robison untermauern, die die Schwächen der Argumente von Behe detailliert darlegt (http://www.talkorigins.org/faqs/behe/review.html). Eine weitere hervorragende Widerlegung von H. Allen Orr findet sich unter http://bostonreview.net/br21.6/orr.html. Siehe auch http://bostonreview.net/evolution.html für mehrere Artikel über Intelligent Design und irreduzible Komplexität von Orr, Russell F. Doolittle, Douglas J. Futuyma, Richard Dawkins, Michael Behe, Phillip Johnson und David Berlinski sowie anderen.
Viele Menschen auf Talk.Origins (T.O.) und anderswo haben die konzeptionellen Gründe dargelegt, warum die darwinistische Evolution irreduzibel komplexe (IC) Systeme produzieren kann. Ich versuche hier, einige Beweise vorzulegen, um diese Konzepte zu stützen. In Robisons FAQ erwähnte er, dass er nicht wisse, ob das Komplementsystem ein evolutionär variables Kaskadensystem zeige. Solche Variabilität über phylogenetische Linien hinweg würde die zentrale Fehlschluss hinter Behes Konzept der irreduziblen Komplexität illustrieren. Dieser Fehlschluss resultiert aus der Unfähigkeit zu erkennen, dass das, was jetzt als irreduzibel komplex erscheint, einfach das Ergebnis der darwinistischen Evolution ist, bei der Merkmale, die einst nur nützlich waren, jetzt essenziell geworden sind. Unten versuche ich, Robisons Punkt zu adressieren und zu zeigen, wie das Komplementsystem Behes Test scheitert.
Übrigens, ein Punkt, den Robison in seinem FAQ machte, war eine Widerlegung der Behauptung von Behe, dass Antikörper nicht eigenständig funktionieren und daher IC sind. Behe liegt zumindest dann falsch, insofern Antikörper Funktionen haben, die über die Rolle hinausgehen, die sie in der Komplementkaskade spielen. Es gibt sogar einige Hinweise darauf, dass Antikörper eine Funktion außerhalb ihrer konventionellen Rolle als „molekulares Etikett" haben. Ich werde hier nicht ins Detail gehen, aber ein gutes Beispiel dafür ist die Tatsache, dass Antikörper katalytisch wirken können. Das heißt, sie können nicht nur als molekulare Etiketten auf Pathogenen, Tumoren oder viral infizierten Zellen fungieren, sondern sie können enzymatische Funktionen haben, die völlig unabhängig von ihren Hauptimmunologischen Rollen sind (siehe zum Beispiel Barbas1).
Zunächst eine kurze Übersicht über das Komplementsystem2
Im Jahr 1899 entdeckte der belgische Jules Bordet, dass Serum zwei Komponenten enthält: eine hitzestabile, die die Aktivität der Agglutination und Fällung aufweist und später von Paul Ehrlich als Antikörper identifiziert wurde, sowie eine hitzelabile Komponente, die für die bakterielle Lyse verantwortlich ist. Erst in den 1950er Jahren wurde jedoch das erste System hitzelabiler Serumproteine (seitdem wurden mehr als 30 identifiziert) aufgeklärt, deren Funktion darin besteht, die antibakterielle Wirkung der Antikörper zu ergänzen. Noch früher, obwohl schlecht verstanden, wurde die Rolle der hitzelabilen Serumproteine erstmals bei frühen Versuchen mit Bluttransfusionen festgestellt, und diese Beobachtungen können uns dabei helfen zu verstehen, wie das Komplementsystem funktioniert.
Die meisten Menschen wissen, dass einer der wichtigsten Bestandteile unseres Immunsystems eine Art Protein namens Antikörper ist. Eine spezialisierte Immunzelle, die B-Zelle, produziert Antikörper. B-Zellen sind Lymphozyten, die eine Schlüsselrolle bei der Beseitigung von Infektionen spielen. Vor allem entwickeln sie sich zu einer Art Zelle, die als Plasmazelle bekannt ist und große Mengen an Antikörpern produziert, obwohl sie auch eine Rolle als antigenpräsentierende Zellen und bei der Rekrutierung von T-Zellen spielen. Unsere Antikörper werden in der Regel so hergestellt, dass sie eindringende Krankheitserreger erkennen, und sind daher für uns nicht schädlich. Manchmal können Antikörper jedoch Schaden anrichten, und Kliniker bezeichnen eine Möglichkeit, wie dies geschehen kann, als hämolytische Transfusionsreaktion, die oft tödlich verläuft. Der häufigste Fall, der heutzutage selten ist, ist die Transfusion von Blutgruppe A in einen Empfänger mit Blutgruppe O. Der Empfänger mit Blutgruppe O bildet Antikörper gegen die roten Blutkörperchen des Spenders mit Blutgruppe A. Kurz nach der Transfusion entwickelt der Empfänger Schüttelfrost, Fieber, Nesselsucht und unkontrollierte Blutungen. Das Plasma des Empfängers ist rosa, aufgrund der Austretung von Hämoglobin aus den roten Blutkörperchen des Spenders. Das Blut ist an Gerinnungsfaktoren verarmt, weil das Gerinnungssystem aktiviert wurde und Blutgerinnsel sich diffus innerhalb der Zirkulation bilden.
Es stellt sich jedoch heraus, dass die Bindung von Antikörpern an Typ-A-Erythrozyten diese nicht schädigt. Wenn wir in Salzlösung suspendierte Erythrozyten mit reinem Antikörper mischen, verklumpen die Zellen, geben jedoch kein Hämoglobin ab. Wenn Blutplasma zu den verklumpten Zellen hinzugefügt wird, lysieren die Zellen, d. h. sie platzen auf, und wenn wir das Plasma einem Empfänger (in einem Tiermodell) verabreichen, reproduziert es die anderen Merkmale einer hämolytischen Transfusionsreaktion. Offensichtlich führt etwas im Plasma diese Effekte aus, nicht der Antikörper. Diese Aktivität wurde als „Komplement" bezeichnet, da sie die Wirkung des Antikörpers ergänzt. Wir wissen heute viel über die Zusammensetzung und Funktion dieser wichtigen Immunfunktion und haben drei Wege identifiziert, auf denen Komplement eindringende Krankheitserreger töten kann.
Aktiviertes Komplementprotein löst eine Kaskade enzymatischer Reaktionen aus, die schließlich zu einem großen Loch führt, das buchstäblich durch die Membran des eindringenden Mikroorganismus oder bei der hämolytischen Transfusionsreaktion in roten Blutkörperchen gebohrt wird. Unsere eigenen Zellen sind vor spontan aktivierten Komplementwegen geschützt (siehe unten), da sie andere Proteine besitzen, die die Komplementkaskade inaktivieren. Nicht überraschend haben einige Mikroorganismen eine Verteidigungsstrategie entwickelt, die das Vorhandensein ihrer eigenen Kaskadeninhibitoren beinhaltet. Die meisten jedoch verfügen nicht über diese Verteidigung, und das Protein-Komplex, der den Mikroorganismus durchbohrt, namens Perforin, erzeugt ein großes Poren in der Membran des Pathogens. Dieser Verlust der Membranintegrität zerstört die Fähigkeit des Pathogens, die Konzentration von Salzen innerhalb seiner selbst zu kontrollieren und tötet es somit.
Unsere komplementären Effektorfunktionen dienen als Arme sowohl des adaptiven als auch des angeborenen Immunsystems. Das angeborene Immunsystem kommt in allen mehrzelligen Organismen vor, während die spezifische (oder adaptive) Immunität nur bei Wirbeltieren anzutreffen ist, mit Ausnahme der Agnather, oder kieferlosen Fische. Antigenrezeptoren im angeborenen System sind germlinell kodiert und erkennen breit homologe Strukturen (meist essenziell), die von Gruppen von Mikroben geteilt werden. Das adaptive Immunsystem entwickelt hingegen sein Antigenrezeptor-Repertoire durch somatische Mutationen, und viele der von diesen Antigenrezeptoren erkannten Determinanten sind idiosynkratisch (meist nicht-essenziell und daher veränderlich) für einen bestimmten Mikroorganismus. Das angeborene Immunsystem entwickelt im Gegensatz zur spezifischen Immunität kein Gedächtnis. Beide Systeme besitzen die Fähigkeit, das Selbst zu unterscheiden, doch ist das spezifische Immunsystem in dieser Hinsicht unvollkommen, was zu Autoimmunität führt.
Die drei Komplementwege sind Waffen, die ein Organismus einsetzen kann, um sich vor Krankheitserregern zu schützen, Tumore zu zerstören und viral infizierte Zellen abzutöten. Sie werden als Lektin-, alternativer und klassischer Weg bezeichnet (bei einigen Organismen gibt es einige geringfügige Variationen). Der Lektinweg, manchmal auch als lytischer Weg bezeichnet, und der alternative Weg gehören zur angeborenen Immunantwort und gelten daher als evolutionär älter als der klassische Weg (siehe unten). Der Lektinweg wird durch Komplementrezeptoren eingeleitet, die aus zirkulierenden Serum-Lektinen bestehen (Lektine sind Proteine, die an Zuckerreste binden); diese binden an Moleküle auf der Oberfläche von Krankheitserregern, die Mannose-Reste enthalten (z. B. Mannan), während der alternative Weg durch Komplement eingeleitet wird, das an die Oberfläche eines Krankheitserregers bindet und spontan aktiviert wird. Der dritte Weg, der aus historischen Gründen als klassischer Weg bezeichnet wird (er wurde zuerst entdeckt), wird durch die Antikörper-Opsonisierung von Krankheitserregern aktiviert (Bindung von Antikörpern an spezifische Epitope von Oberflächenproteinen, z. B. Antigene) und gehört daher zur adaptiven Immunität.
Jeder der drei Wege hat eigene frühe Ereignisse, einschließlich der Bindung von Rezeptoren und eines einzigartigen Satzes von Komponenten des Komplementsystems (siehe unten), die eine enzymatische Kaskade einleiten. Alle drei Wege konvergieren bei einer Enzymaktivität, die als C3-Konvertase bezeichnet wird. Bei Wirbeltieren ist das C3-Konvertase-Enzym sowohl im Lektin- als auch im klassischen Weg ein Protein namens C2b, während es im alternativen Weg eine funktionell und strukturell homologe Kombination aus zwei Proteinen, C3b und dem Plasmafaktor B, darstellt. Diese Kombination wird ihrerseits durch den Plasma-Protease-Faktor D aktiviert. An diesem Punkt haben alle drei Wege ähnliche Effektorfunktionen. Im Folgenden ist ein Schema der wichtigsten Akteure in allen drei Komplementaktivierungswegen von Wirbeltieren* (bitte beachten Sie: leider wurden viele Proteine in der Reihenfolge ihrer Entdeckung nummeriert, nicht in ihrer jeweiligen Reihenfolge in den Wegen).
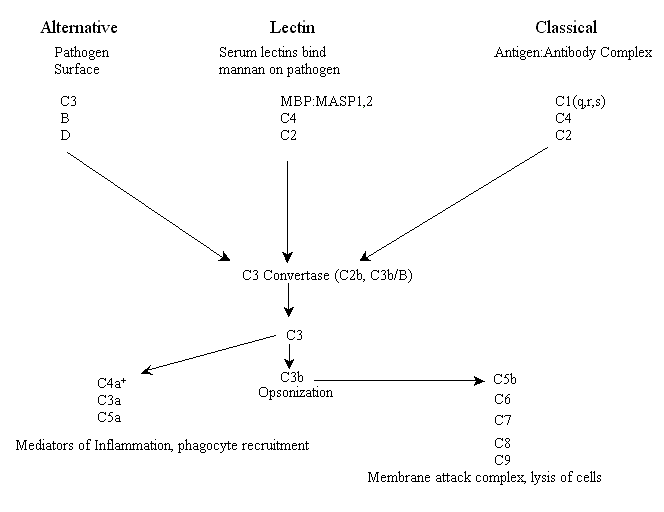
+C4a ist ein Produkt der Spaltung von C4 durch MASP oder C1s, nicht durch C3, und dient als Vermittler der Entzündung.
|
Zusammenfassung der an der Komplementkaskade beteiligten Proteine |
|
|
Bindung an Ag:Ab-Komplexe |
C1q |
|
Aktivierung von Enzymen |
C1r, C1s, C2b, Bd, D, MASP1,2 |
|
Membranbindende Opsonine |
C4b, C3b, MBP |
|
Entzündungsmediatoren |
C5a, C3a, C4a |
|
Membranangriff |
C5b, C6, C7, C8, C9 |
|
Komplementrezeptoren |
CR1, CR2, CR3, CR4, C1qR |
|
Komplementregulatorische Proteine |
C1INH, C4bp, CR1, MCP, DAF, H, I, P, CD59 |
*Angelehnt an Janeway & Travers Immunobiology, 1996; Current Biology Ltd/Garland Publishing Inc.
Evolutionäre Geschichte der Komplementwege
Alle untersuchten Wirbeltiere mit Ausnahme der Agnathen teilen das menschliche Komplementsystem. Je weiter wir phylogenetisch entfernt sind, desto unterschiedlicher sind sowohl das Immunsystem als auch seine Effektoren, einschließlich der Komplementsysteme. Drei davon wurden untersucht: der Lachsfisch (eine Agnathen), die Ascidien-Gruppe der Urochordaten und die Seesterne. Es gab viel Debatten über die evolutionäre Geschichte der Komplementwege. Es gibt einige Hinweise darauf, dass bei Wirbeltieren der alternative Weg dem klassischen Weg vorausgeht, basierend auf Sequenzvergleichen des ersten Komplementkomponenten, C1q3. Allerdings deutet andere Evidenz darauf hin, dass der alternative Weg älter ist als der klassische Weg, basierend auf phylogenetischen Vergleichen von C3a, C3b und C5a4. Darüber hinaus besteht das Komplementsystem des Lachsfisches nur aus dem alternativen und dem Lektinweg5. Kürzlich wurde gezeigt, dass ein Urochordat Gene besitzt, die den Wirbeltiergenen C3, C4 & C5 entsprechen, aber nicht C26. Weiterhin zeigten Ji et al7 und Nonaka, M et al8, dass der Lektinweg evolutionär der älteste der drei Wege ist, indem sie nachwiesen, dass MASP in einem Urochordat, der Ascidie Halocynthia roretzi (Japanische Seesquirt), vorkommt. Es wird angenommen, dass ein Großteil des Wirbeltier-Komplementsystems aus der Duplikation von Genen hervorgegangen ist, die C3/C4/C5, Bf/C2, C1s/C1r/MASP-1/MASP-2 und C6/C7/C8/C9 Moleküle kodieren9
Ist der Komplementweg irreduzibel komplex?
Einer der Behauptungen von Behe ist, dass Enzymkaskaden wie die Aktivierung des Komplementsystems irreduzibel komplex sind, da jeder neue Schritt in der Kaskade "sowohl ein Proenzym als auch ein aktivierendes Enzym erfordern würde, um das Proenzym zur richtigen Zeit und am richtigen Ort zu aktivieren". Er weist unter anderem auf das Komplementsystem hin, das als irreduzibel komplex erscheint. Und so scheint es. Zum Beispiel stützen sich alle drei Kaskaden auf die Aktivität des C3-Konvertasen, um das Komplement zu aktivieren und dem Organismus zu helfen, sich vor Pathogenen zu schützen. Menschen, die C3 fehlen, haben keine funktionsfähigen Komplementwege und leiden unter einer erhöhten Anfälligkeit für pyogene Infektionen durch pathogene Bakterien wie Haemophilus influenzae und Streptococcus pneumoniae unter anderem. Es scheint dann, dass die Wirbeltierwege nicht auf darwinistische Weise von früheren Lektinwegen evolviert sein könnten, da sie von der Aktivität des C3-Konvertasen abhängig sind. Aber das liegt daran, dass Behe und andere nicht erkennen, dass die aktuellen Akteure in der Kaskade nicht immer vorhanden waren.
Es gibt zwei allgemeine Linien von Beweisen, die darauf hindeuten, dass das Komplementsystem bei Säugetieren zu seinem heutigen Status evolviert ist. Erstens sehen wir Komplementsysteme bei Wirbellosen, die einen ganzen Arm des Säugetier-Systems vermissen. Zweitens sind die Proteine der Urochordaten- und Wirbeltier-Systeme homolog. Insbesondere gibt es Homologien zwischen Proteasen, die auf Genduplikation und anschließende Spezialisierung hindeuten, wie oben erwähnt. Die Situation ähnelt der Gerinnungskaskade, bei der Homologien beobachtet werden, die durch Genduplikation erklärt werden können. Organismen, die phylogenetisch weit von Menschen entfernt sind und somit eine Linie von Tieren darstellen, die evolutionär älter als heutige Wirbeltiere sind, teilen dennoch den Lektinweg mit uns. Dies ist eines von sehr vielen Stücken von Beweisen für gemeinsame Abstammung. Ein weiterer ist die Tatsache, dass Wirbeltiere einen gesamten Weg besitzen, der bei Vertretern evolutionär älterer Organismen fehlt.
Lassen Sie uns genauer auf den Lektinweg, den Komplementweg, der von Urochordaten und Wirbeltieren gemeinsam genutzt wird, eingehen. Er wird durch die Bindung von Mannan-bindendem Protein (MBP) und dem MBP-assoziierten Serinprotease (MASP) eingeleitet, das als Proenzym vorliegt. MBP erkennt N-Acetylglucosamin-Reste an Oberflächenproteinen von Pathogenen und aktiviert dadurch MASP. Die aktivierten MASP spaltet dann C4 und C2 proteolytisch. C2 wird gespalten, um C2b und C2a zu bilden. C2b wird auch als C3-Konvertase bezeichnet, die enzymatische Aktivität, die allen drei Wegen gemeinsam ist. Die C3-Konvertase ist für die Komplementwege von Säugetieren von entscheidender Bedeutung, sodass Mutationen in C2 (und C3) Krankheitszustände bei Ratten, Mäusen, Meerschweinchen, Primaten und Menschen hervorrufen können (siehe Übersicht in 10-12). Das Löschen des C3-Gens führt zur Auslöschung der Komplementwege, wie bei Knockout-Mäusen beobachtet15.
Das Interessante am Seespinn C3-Protein ist, dass es keinen C3-Konvertase-Spaltungsstellen enthält, die in allen bisher bekannten Wirbeltier-C3-Sequenzen vorkommen7. Dies bedeutet, dass ein Enzym, das sich von der Wirbeltier-C3-Konvertase (C2b) unterscheidet, das C3-Protein von H. roretzi aktiviert. Tatsächlich besitzen Urochordaten kein C2-Gen6. Ein Kandidat für die Urochordaten-C3-Konvertase ist MASP. MASP scheint durch Sequenz-Homologie mit den C1-Enzymen des klassischen Weges verwandt zu sein, von denen eines, C1s, C4 spaltet13, 14, was auf eine Quelle für die Wirbeltier-C1-Genen durch Gen-Duplikationen hindeutet. Es sind zwei MASP-Enzyme bekannt, die als aktivierende Enzyme in Komplementwegen fungieren. MASP-1 bindet an MBP (Hinweis: MBP wird manchmal als MBL für Mannan Binding Lectin bezeichnet). Diese Bindung aktiviert MASP-1, welches dann C4 und C2 spaltet, wie oben erwähnt. Es wurde kürzlich postuliert, dass MASP-1 auch eine zweite Serin-Protease, genannt MASP-2, aktiviert, die ein Wirbeltier-C3-Konvertase-Enzym im Lektin-Weg produziert13. Ein drittes MASP-Enzym, MASP-3, wurde identifiziert und soll die Aktivität der anderen MASPs regulieren15.
Kann MASP selbst C3 aktivieren, indem es als C3-Konvertase fungiert? Ja. Säugetier-MASP kann C3 direkt aktivieren, tut dies jedoch schlecht16, 17, und Säugetiere, die C3 nicht umwandeln können, leiden unter einer Vielzahl von Krankheiten10-12. Ascidien-MASP hingegen ist bekannt für C3-Konvertase-Aktivität18. Da Urochordaten C2 und somit C2b fehlen, aber MASP besitzen, scheinen sie ein verkürztes, aber offensichtlich wirksames Komplementsystem zu haben, das den Bedarf an der C3-Konvertase C2b umgeht. Während die klassischen Wege der Wirbeltiere irreduzibel komplex sind, da der gesamte Weg von mindestens einem Enzym abhängt, ist klar, dass evolutionär ältere Organismen einen funktionellen Weg besitzen, auch in Abwesenheit dieses Enzyms. Im Wirbeltier-Lektinweg übernahm die C3-Konvertase C2b die Funktion der wirbellosen MASP, sodass das Wirbeltier-Komplementsystem nun kritisch von der Aktivität von C2b abhängt.
Dies deutet darauf hin, dass sich die Wirbeltiere, als sie entstanden und ihre eigenen Komplementsysteme entwickelten, auf den Kaskadenmechanismus der Wirbellosen stützten, jedoch einen eigenen Bestandteil hinzufügten, C2 (unter anderem). Dies ermöglichte es den Wirbeltieren, die vollen Vorteile ihrer neuen Art von Immunität, der adaptiven Immunität, zu nutzen. Sie behielten den lytischen Weg bei, etablierten jedoch den klassischen Komplementweg, indem sie das Modell der Wirbellosen übernahmen. Im Laufe der Zeit wurde der klassische Weg der Wirbeltiere jedoch kritisch abhängig vom neuen Bestandteil C2, den frühere Wege nicht benötigten. Die Wirbeltiere entwickelten zudem den alternativen Weg, indem sie erneut auf den Weg der Wirbellosen zurückgriffen.
Dies muss wiederholt werden: Urochordaten besitzen ein funktionelles Komplementsystem, fehlen jedoch ein Bestandteil der Kaskade, den C3-Konvertase, der in derselben Kaskade bei Wirbeltieren essenziell ist. Erinnern Sie sich an das Zitat von Dr. Behe am Anfang dieses Essays: „Ein irreduzibel komplexes System kann nicht schrittweise durch geringfügige, aufeinanderfolgende Modifikationen eines Vorläufersystems entstehen, da jeder Vorläufer eines irreduzibel komplexen Systems per Definition nicht funktionsfähig ist". Dieses Beispiel einer irreduzibel komplexen Komplementkaskade, die eindeutig das Ergebnis darwinischer Evolution ist, widerlegt die Behauptung von Dr. Behe.
Zusammenfassend gibt es Probleme mit Behes Behauptung, dass das Komplement ein „irreduzibel komplexes System" sei oder dass es unwahrscheinlich sei, durch Evolution entstanden zu sein. Obwohl es hochgradig ausgearbeitet ist, bedeutet das Entfernen einiger Teile sicher nicht, dass das System „aufhört zu funktionieren". Das Vorkommen einfacherer Systeme in phylogenetisch älteren Organismen deutet auf einen evolutionären Pfad hin, und das Vorkommen zahlreicher Homologien zwischen den Genen deutet auf einen Mechanismus für seine Evolution hin. Dies demonstriert den Punkt, den Keith Robison, H. Allen Orr, Russell Doolittle, Douglas Futuyma und eine Vielzahl von T.O.-Regulären immer wieder versucht haben zu machen. Um Dr. Orr (IIRC) zu paraphrasieren: „Darwinische Evolution kann irreduzible Komplexität leicht hervorzubringen: alles, was erforderlich ist, ist, dass Teile, die einst nur vorteilhaft waren, aufgrund späterer Veränderungen zu essentiellen werden."19
Vielen Dank an George Acton für seine fundierten Vorschläge und seine Kritik an einem Entwurf dieses Essays.
Referenzen
3. Dodd, A.W., & F. Petry, Behring Inst Mitt 1993 (93):87-102.
4. Farries, TC, Steuer KL und JP Atkinson, Immunol Today 1990 (3):78-80.
6. Smith, L.C., Chang, L., Britten, R.J., & E.H. Davidson, J., Immunol. 1996 (156):593-602.
7. Ji, X., Azumi, K., Sasaki, M.,& N. Masaru, Proc. Natl. Acad. Sci USA, 1997 (94):6340-6345.
9. Zarkadis IK, Mastellos D, Lambris JD, Dev Comp Immunol 2001 Oktober-Dezember;25(8-9):745-62
10. Sakamoto M, Fujisawa Y, Nishioka K Nutrition 1998 (4):391-8.
11. Carroll MC Annu Rev Immunol 1998 (16):545-68
12. Frank M.M., J Clin Immunol 1995 (6 Suppl):113S-121S.
13. Vorup-Jensen T, Jensenius JC, Thiel S. Immunobiologie 1998 Aug;199(2):348-57
14. Matsushita M, Endo Y, Fujita T. Immunobiology 1998 Aug;199(2):340-7
16. Ogata, R.T., Low, P. J. & M. Kawakami, J. Immunol. 1995 (154):2351-2357
17. Matsushita, M. & T. Fujita Immunobiologie 1995 (194):443-448
18. Nonaka M, Azumi K. Dev Comp Immunol 1999 Jun-Jul;23(4-5):421-7
19. Siehe http://bostonreview.net/br21.6/orr.html.
[Zurück zu den Häufig gestellten Fragen zur irreduziblen Komplexität]