Zu lesen in Verbindung mit Douglas Theobalds FAQ 29+ Beweise für Makroevolution.
Diese FAQ behandelt folgende Themen:
-
Was makroevolution und mikroevolution bedeuten
-
Wie die Begriffe verwendet werden und wie sie entstanden sind
-
Verwirrungen in der wissenschaftlichen Literatur über die Begriffe
-
Eine philosophische Diskussion darüber, ob makroevolution auf mikroevolution zurückführbar ist oder ob sie als ein separater Prozess in der Evolution steht
-
Ob es Barrieren gibt, die verhindern, dass mikroevolution, die Kreationisten akzeptieren, zu makroevolution wird, die sie ablehnen
-
Ob die Idee der makroevolution widerlegbar ist und ob spezifische Darstellungen der makroevolution widerlegbar sind.
Antievolutionisten argumentieren so laut gegen die Makroevolution, dass einige Menschen glauben, der Begriff sei erfunden worden, um die Evolution abzutun. Aber das ist nicht wahr; Wissenschaftler verwenden nicht nur diese Begriffe, sie haben zudem ein ausgearbeitetes Set von Modellen und Ideen dazu, die Antievolutionisten natürlich übergehen oder als Probleme für die Evolutionsbiologie darstellen.
Eine spätere Version wird einen Abschnitt hinzufügen, der beschreibt, wie Kreationisten, wenn sie mit unbestreitbaren Beweisen für die Makroevolution konfrontiert werden, „die Torhürten verschieben". Für jetzt siehe die Schwester-FAQ von Douglas Theobald.
Der Leser wird eingeladen, den Abschnitt über die Reduktion zu überspringen. Dies ist eine weitgehend philosophische Diskussion, die aufgenommen wurde, weil es sich um eine Debatte innerhalb der wissenschaftlichen Gemeinschaft handelt. Sie hat keinen Einfluss auf die Tatsache der evolutionären Entwicklung über die Art hinaus (das heißt, auf die Artbildung, die gemeinsame Abstammung und das Muster im phylogenetischen Fossilbericht). Doch sie ist oft Gegenstand heftiger Debatten in Foren, die Evolution im Kontext des Kreationismus diskutieren.
Wie Humpty Dumpty zu Alice sagte:
Es gibt für dich Ruhm!
'Ich weiß nicht, was du mit "Ruhm" meinst,' sagte Alice.
Humpty Dumpty lächelte verächtlich. 'Natürlich nicht – bis ich es dir sage. Ich meinte: "Es gibt für dich ein schönes, entscheidendes Argument!"'
'Aber "Ruhm" bedeutet nicht "ein schönes, entscheidendes Argument",' wandte Alice ein.
'Wenn ich ein Wort benutze,' sagte Humpty Dumpty in einem ziemlich verächtlichen Ton, 'dann bedeutet es genau das, was ich es sein lassen will – nicht mehr und nicht weniger.'
'Die Frage ist,' sagte Alice, 'ob du Wörter dazu bringen kannst, so viele verschiedene Dinge zu bedeuten.'
'Die Frage ist,' sagte Humpty Dumpty, 'wer der Herrscher sein soll – das ist alles.'
Wörter sind nicht die Herrscherin der Wissenschaft; die Wissenschaft ist es, oder sollte es sein, über ihre Wörter. Aber wir können untersuchen, wie Wissenschaftler ihre Wörter verwenden und ob sie sie konsistent einsetzen. Und nachdem wir das getan haben, können wir untersuchen, ob andere, die keine Wissenschaftler sind, zu viel hineininterpretieren oder sie auf eine völlig andere Weise verwenden.
Inhalt
- Was ist Makroevolution?
- Die Geschichte des Konzepts der Makroevolution
- Verwirrungen
- Ist Makroevolution auf Mikroevolution reduzierbar?
- Hindernisse für Makroevolution
- Widerlegung der Makroevolution
- Fazit
- Anmerkungen
- Literatur
Was ist Makroevolution?
Zuerst müssen wir die Definitionen korrekt festlegen. Die folgenden Begriffe werden definiert: Makroevolution, Mikroevolution, Kladogenese, Anagenese, Theorie der unterbrochenen Gleichgewichte, phyletischer Gradualismus
Kreationisten behaupten oft, dass "Makroevolution" nicht bewiesen sei, auch wenn "Mikroevolution" es ist, und damit meinen sie anscheinend, dass alles, was an Evolution beobachtet wird, Mikroevolution ist, der Rest aber Makroevolution. Bei der Aufstellung dieser Behauptungen missbrauchen sie authentische wissenschaftliche Begriffe; das heißt, sie verwenden eine nicht-standardisierte Definition, mit der sie versuchen, den Anschein zu erwecken, die Wissenschaft sage etwas anderes als es tatsächlich tut. Befürworter der Evolution sagen oft, dass Kreationisten die Begriffe erfunden haben. Das ist falsch. Sowohl makroevolution als auch mikroevolution sind legitime wissenschaftliche Begriffe, die eine Geschichte von sich wandelnden Bedeutungen haben, die ohnehin nicht den Kreationismus stützen können.
In der Wissenschaft bedeutet „makro-" am Anfang eines Wortes einfach „groß" und „mikro-" am Anfang eines Wortes einfach „klein" (beide aus den griechischen Wörtern). Zum Beispiel bedeutet „Makrofauna" große Tiere, die mit bloßem Auge beobachtet werden können, während „Mikrofauna" kleine Tiere bezeichnet, die möglicherweise beobachtbar oder möglicherweise nicht ohne ein Mikroskop sind. Etwas kann „makro" sein, indem es einfach größer ist, oder es kann einen Übergang geben, der es zu etwas ganz Distinktem macht.
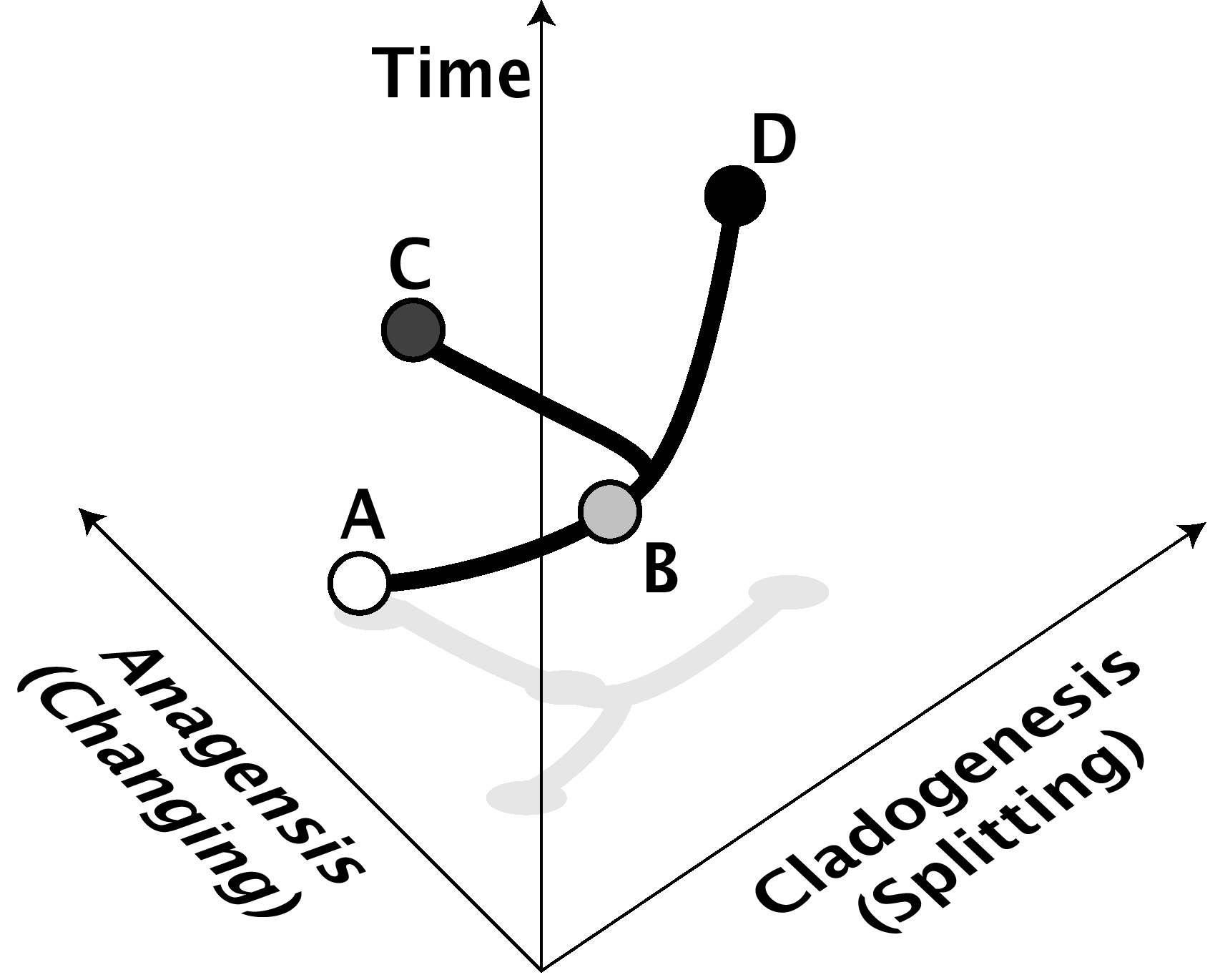
In der heutigen Evolutionsbiologie wird makroevolution für jede evolutionäre Veränderung verwendet, die auf oder über der Artenebene stattfindet. Es bedeutet mindestens die Aufspaltung einer Art in zwei (Artbildung, oder kladogenese, vom griechischen für "die Entstehung eines Zweigs", siehe Abb. 1) oder die Veränderung einer Art im Laufe der Zeit in eine andere (anagenetische Artbildung, heutzutage nicht allgemein akzeptiert [Hinweis 1]). Jede Veränderung, die auf höheren Ebenen auftritt, wie die Evolution neuer Familien, Stämme oder Gattungen, ist ebenfalls daher Makroevolution, aber der Begriff ist nicht auf diese höheren Ebenen beschränkt. Er bedeutet oft auch langfristige Trends oder Verzerrungen in der Evolution höherer taxonomischer Ebenen.
Mikroevolution bezeichnet jede evolutionäre Veränderung unterhalb der Art-Ebene und bezieht sich auf Änderungen der Frequenz innerhalb einer Population oder einer Art ihrer Allele (alternative Gene) und deren Auswirkungen auf die Form, oder das Phänotyp, der Organismen, die diese Population oder Art bilden. Sie kann sich auch auf Veränderungen innerhalb von Arten beziehen, die nicht genetisch sind.
 d
d |
Abbildung 1: Anagenese und Kladogenese. In diesem Beispiel verändert sich die Art A anagenetisch über die Zeit zur Art B, während sich die Art B kladogenetisch über die Zeit durch Aufspaltung in die Arten C und D verändert, von denen keine sehr unterschiedlich von B oder voneinander sind. Die anagenetische Achse repräsentiert Formveränderung, entweder genetisch oder phänotypisch. Die kladogenetische Achse repräsentiert Isolation der Arten voneinander (zum Beispiel reproduktive Isolation). Natürlich können Kladogenese und Anagenese oft auch Hand in Hand gehen. Anagenese wird von den meisten Wissenschaftlern nicht als „echte" Artbildung betrachtet, obwohl sie im Fossilbericht von einem kladogenetischen Ereignis nicht zu unterscheiden ist. |
Ein anderer Weg, den Unterschied zu beschreiben, ist, dass Makroevolution die Evolution zwischen Arten ist und Mikroevolution die Evolution innerhalb einer Art. Manchmal wird Makroevolution als „supraspezifische Evolution" bezeichnet (Rensch 1959, siehe Hennig 1966: 223-225).
Es gibt verschiedene Ansichten über die Dynamik der Makroevolution. Punctuated Equilibria sind Muster des Wandels, die Stase anzeigen, also lange Zeiträume, in denen Arten sehr wenig Veränderung zeigen. Es gibt mehrere Hypothesen, die versuchen, Stase zu erklären. Der aktuelle Konsens unter Paläontologen ist, dass große Populationen durch natürliche Selektion oder genetische Drift gegen evolutionäre Veränderungen abgeschirmt werden. Evolutionäre Veränderungen werden leichter, wenn Populationen in kleinere Demen aufspalten. Diese Veränderung kann „eingefroren" werden, wenn die Teilpopulationen reproduktive Isolation entwickeln und zu eigenen Arten werden. Deshalb wird Veränderung mit Kladogenese in Verbindung gebracht. Phyletic gradualism besagt, dass Arten im Laufe ihrer Geschichte weiterhin an neue Herausforderungen angepasst werden (siehe Abb. 1). Species selection und species sorting Theorien gehen davon aus, dass makroevolutionäre Prozesse ablaufen, die es wahrscheinlicher oder unwahrscheinlicher machen, dass bestimmte Arten sehr lange existieren, bevor sie aussterben, in einer Art Parallel zu dem, was bei Genen in der Mikroevolution passiert.
Die Geschichte des Konzepts der Makroevolution
Wie sind die Begriffe in den wissenschaftlichen Gebrauch eingegangen, und was ist seitdem mit ihnen geschehen?
In der „modernen Synthese" des Neodarwinismus, die sich in der Zeit von 1930 bis 1950 durch die Versöhnung der Evolution durch natürliche Selektion und der modernen Genetik entwickelte, wird Makroevolution als die kombinierten Effekte von Mikroevolutionsprozessen betrachtet.
Die Begriffe makroevolution und mikroevolution wurden erstmals 1927 vom russischen Entomologen Iuri'i Filipchenko (oder Philipchenko, je nach Transkription) in seiner deutschsprachigen Arbeit Variabilität und Variation geprägt, die einen frühen Versuch darstellte, Mendelsche Genetik und Evolution in Einklang zu bringen. Filipchenko war ein Evolutionist, doch da er während der Zeit schrieb, in der es schien, als habe der Mendelismus den Darwinismus überflüssig gemacht, die sogenannte „Eclipse of Darwinism" (Bowler 1983), war er kein Darwinist, sondern ein Orthogenetiker (er glaubte, die Evolution habe eine Richtung). Darüber hinaus hatten russische Biologen dieser Zeit eine Geschichte des Ablehnens von Darwins malthusianischem Mechanismus der Evolution durch Konkurrenz (Todes 1989).
In Dobzhansky's grundlegenden Werk der modernen Synthese, Genetics and the Origin of Species, begann er damit, festzustellen, dass wir „auf der gegenwärtigen Ebene des Wissens reluktant gezwungen sind, ein Gleichheitszeichen zwischen den Mechanismen der Makro- und der Mikroevolution zu setzen" (1937: 12), wodurch er diese Begriffe in die englischsprachige biologische Gemeinschaft einführte (Alexandrov 1994). Dobzhansky war Filipchenkos Schüler und betrachtete ihn als seinen Mentor. In der Wissenschaft wie in allen akademischen Disziplinen ist es schwierig, eine zentrale Lehre seiner Lehrer aufgrund familiärer Loyalität zu bestreiten, und Dobzhansky, der mit diesem Buch effektiv die moderne darwinistische Synthese einleitete, fand es unangenehm, die Ansichten seines Lehrers bestreiten zu müssen (Burian 1994).
Der Begriff geriet in begrenztem Misskredit, als er von Autoren wie dem Genetiker Richard Goldschmidt (1940) und dem Paläontologen Otto Schindewolf übernommen wurde, um ihre orthogenetischen Theorien zu beschreiben. Infolgedessen bedienten sich, außer Dobzhansky, Bernhardt Rensch und Ernst Mayr, sehr wenige neodarwinistische Autoren des Begriffs und bevorzugten es stattdessen, von Evolution als Veränderungen der Allelfrequenzen ohne Erwähnung der Ebene der Veränderungen (oberhalb oder unterhalb der Artenebene) zu sprechen. Diejenigen, die dies taten, arbeiteten im Allgemeinen innerhalb der kontinental-europäischen Traditionen (wie Dobzhansky, Mayr, Rensch, Goldschmidt und Schindewolf), während diejenigen, die dies nicht taten, im Allgemeinen innerhalb der angelsächsischen Tradition arbeiteten (wie John Maynard Smith und Richard Dawkins). Daher wird die Verwendung des Begriffs „makroevolution" manchmal fälschlicherweise als Leitschritt verwendet, um festzustellen, ob der Autor „richtig" neodarwinistisch ist oder nicht (Eldredge 1995: 126-127).
Der Begriff wurde von einer Reihe vor allem paläontologischer Autoren wie Steven Stanley (1979), Stephen Jay Gould und Niles Eldredge, den Autoren der Theorie des unterbrochenen Gleichgewichts (siehe Eldredge 1995), wiederbelebt, die argumentierten, dass etwas anderes als innerhalb der Art ablaufende Prozesse die Makroevolution verursachen, obwohl sie die Ansicht ablehnen, dass die Evolution progressiv ist. Viele Paläontologen haben behauptet, dass das, was in der Evolution über das Artniveau hinaus geschieht, auf Prozesse zurückzuführen ist, die über das Niveau der Populationen hinaus wirken – zum Beispiel die Vorstellung der Artselektion (die Idee, dass Arten selbst ähnlich selektiert werden, wie Allele innerhalb von Populationen selektiert werden, siehe Grantham 1995, Rice 1995 und Stidd und Wade 1995 für Übersichten und Diskussionen).
Die Vorstellung, dass die Entstehung höherer Taxa wie Gattungen etwas Besonderes erfordert, basiert oft auf einem Missverständnis der Art und Weise, in der neue Linien entstehen. Die beiden Arten, die die Ursprünge der Hund- und Katzenlinien bilden, unterschieden sich wahrscheinlich nur sehr wenig von ihrer gemeinsamen Vorfahrenart und voneinander. Sobald sie jedoch taxonomisch voneinander isoliert waren, entwickelten sie zunehmend Unterschiede, die sie intern teilten, die aber andere Linien nicht aufwiesen. Dies gilt für alle Linien zurück bis zur ersten eukaryotischen (kernhaltigen) Zelle. Selbst die Veränderungen während der Kambrium-Explosion sind von dieser Art, obwohl einige (z. B. Gould 1989) der Meinung sind, dass die Genome (Genstrukturen) dieser frühen Tiere nicht so streng reguliert waren wie bei modernen Tieren und daher mehr Spielraum für Veränderungen hatten.
Verwirrungen
Ways in which the term "macroevolution" is used by scientists. Some are exact in the way they use it, while others are less exact. These usages are not all the same, and this causes some confusion. Why do scientists not agree on the meaning of their terms?
Die Bedeutung, die modernen Autoren den Begriffen "Makroevolution" und "Mikroevolution" geben, ist oft verwirrend und variiert je nachdem, worüber sie sprechen. Dies gilt insbesondere, wenn über "großräumige" evolutionäre Prozesse gesprochen wird. Zum Beispiel definiert R. L. Carroll in seinem Lehrbuch für Studierende (1997: 10) Mikroevolution als "Phänomene auf der Ebene von Populationen und Arten" und Makroevolution als "evolutionäre Muster, die sich über Millionen und Hunderte von Millionen Jahren erstrecken". Eldredge sagt: "Makroevolution, wie auch immer sie präzise definiert wird, impliziert immer 'großräumige evolutionäre Veränderung' (1989: vii) und spricht in seinem Buch über Makroevolution als grob gleichbedeutend mit der Evolution von Taxa, die einen höheren Rang als die Art haben, wie Gattungen, Ordnungen, Familien und dergleichen. In seinem Buch Evolution definiert Mark Ridley die Begriffe wie folgt (2004: 227):
Die Makroevolution bedeutet Evolution im großen Maßstab und wird hauptsächlich im Fossilbericht untersucht. Sie wird der Mikroevolution gegenübergestellt, die die Evolution über kurze Zeiträume untersucht, wie etwa innerhalb eines menschlichen Lebens oder weniger. Mikroevolution bezieht sich daher auf Veränderungen der Allelfrequenz innerhalb einer Population .... Makroevolutionäre Ereignisse sind viel wahrscheinlicher über Millionen von Jahren. Makroevolution bezieht sich auf Dinge wie die Trends in der Evolution des Pferdes ... oder den Ursprung großer Gruppen, oder Massenaussterben, oder die Kambrium-Explosion .... Artbildung ist die traditionelle Trennlinie zwischen Mikro- und Makroevolution.
Es gibt viele veröffentlichte Arbeiten, die den Begriff in dieser Art der „höheren Kategorie" verwenden; warum ist das so?
Die Wissenschaft ist in ihrer Verwendung von Begriffen nicht immer konsistent; dies ist die Quelle vieler Verwirrungen. Manchmal handelt es sich dabei um Nachlässigkeit, und manchmal liegt es an der Art und Weise, wie sich Begriffe im Laufe der Zeit entwickeln. Wenn Biologen und Paläontologen über Makroevolution im Sinne von „großräumiger" Evolution sprechen, meinen sie streng genommen nur einen Teil der Phänomene, die der Begriff umfasst, aber es ist der interessanteste Teil für diese Spezialisten. Das heißt, sie sprechen über die Muster der Evolution weit über der Artenebene hinaus (Smith 1994).
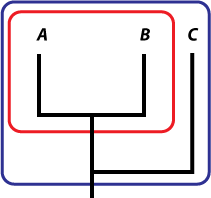
Um ein Muster zu erkennen, müssen Sie mindestens drei Arten vergleichen (Abb. 2). Allein betrachtet bildet Art A keine Muster, und solange die Veränderungen innerhalb dieser Art nicht zu einer neuen Art führen, handelt es sich um Mikroevolution. Wenn sich eine neue Art B von A abzweigt, liegt dann zwar Makroevolution vor, aber keine Muster. Damit ein Muster vorliegt, müssen Sie feststellen können, dass eine Art enger mit einer anderen verwandt ist als mit einer dritten (in diesem Fall, dass A enger mit B verwandt ist als mit C).
 d d |
Abbildung 2: Wenn nur zwei Arten oder höhere Taxa identifiziert werden (rote Menge), gibt es kein Muster. Wenn drei oder mehr (blaue Menge) einbezogen werden, kann man sagen, dass eine Art evolutionär gesehen enger mit einer anderen verwandt ist als mit der dritten – in diesem Fall sind A und B enger miteinander verwandt als mit C, das sich früher vom A/B-Zweig abgespalten hat. |
Die Muster, die Menschen bei der Diskussion der Makroevolution interessieren, betreffen tendenziell sehr viele Arten, entweder als einzelne große Gruppe ("höhere Taxa") oder einzeln. Aus diesem Grund verwenden viele Autoren den Begriff "Makroevolution", um "großräumige Evolution" zu bedeuten. Allerdings ist "großräumig" genau wie bei anagenetischer Artbildung ein willkürlicher und oft subjektiver Begriff, und die objektive Bedeutung der Makroevolution ist Evolution auf oder über der Artenebene [Hinweis 2]. Daher ist Carrolls "Definition" problematisch, trotz seiner Prominenz in diesem Feld, und dieser Art von Verwirrung ist zu vermeiden. Ein vorheriger Versuch von Simpson (1944), "Megaevolution" für großräumige Veränderungen einzuführen, wurde ebenfalls nicht akzeptiert, zum Teil, weil nie ganz klar war, wann "makro" endete und "mega" begann.
Eine wohlüberlegtere Definition ist die von Levinton: "Ich definiere den Prozess der Makroevolution als 'die Summe jener Prozesse, die die Charakterzustandsübergänge erklären, die evolutionäre Unterschiede von höherer taxonomischer Rangordnung diagnostizieren'" (Levinton 2001:2). Hier versucht Levinton, Makroevolution so zu definieren, dass er dem Debattenfeld, über das er schreibt, nicht vorurteilsbehaftet gegenübersteht. Er konzentriert sich auf die Merkmale von Taxa und ist neutral bezüglich der Taxon-Ebene, die involviert ist. Er verneint die Definition auf der "Art-Ebene", weil er glaubt – ich glaube unnötigerweise –, dass dies Makroevolution zur Studie der Artbildung machen würde. Wenn die obige Analyse des "Musters" zutrifft, dann umfasst Makroevolution lediglich die Studie der Artbildung, ist aber kaum darauf beschränkt. Der Umfang der Makroevolution reicht weit über diese Ebene hinaus. Es ist jedoch erwähnenswert, dass die höheren taxonomischen Ebenen nach Linnaeus künstlich sind und von Systematikern aus Bequemlichkeit konstruiert wurden. Schlussfolgerungen über die Evolution, die auf taxonomischen Ebenen wie Gattungen oder Familien beruhen (z. B. die Arbeit von Raup und Sepkoski über das Aussterben, Raup und Sepkoski 1986, Sepkoski 1987, Raup 1991), müssen mit einer Prise Salz genommen werden, da die Taxon-Ebenen bei phylogenetisch entfernten Gruppen nicht die "gleichen" sind, weil sie nicht "natürlich" sind, obwohl sie tatsächlich (wie gezeigt werden wird) gute Surrogate für phylogenetische Vielfalt sein können.
Übrigens hat sich die Erforschung der Artbildung in den letzten Jahren erheblich entwickelt, gestützt durch fundierte theoretische Arbeiten, die nahelegen, dass viele makroevolutionäre Effekte tatsächlich das Ergebnis von Prozessen auf Populationsebene sind (Gavrilets 2003, 2004, Gavrilets und Gravner 1997). Unter Verwendung der Metapher der adaptiven Landschaft – des Feldes aller möglichen Genrekombinationen für eine Population, wofür jedes eine Fitnesswert durch die Umwelt zugewiesen bekommt – haben Gavrilets und Kollegen gezeigt, dass das, was auf Populationsebene geschieht, tatsächlich zu einer Divergenz zwischen ihnen führen kann, dass aber die meisten Male Populationen, die durch Selektion auf eine hohe Fitness gehalten werden, dennoch zufällig „driften" können.
Ist Makroevolution auf Mikroevolution reduzierbar?
Heißes Gerücht: Wissenschaftsphilosophen mögen gerne darüber streiten, ob eine Wissenschaft auf eine andere reduziert werden kann. Viele haben gefragt, ob Makroevolution auf Mikroevolution reduziert werden kann. Das heißt, ob größere Veränderungen in der Evolution „nur die Summe von“ kleinen Veränderungen sind. Wir müssen verstehen, was „Reduktion“ in der Wissenschaftsphilosophie bedeutet, bevor wir beginnen, Menschen als „Reduktionisten“ oder „Holisten“ zu beschuldigen.
Aus philosophischer Sicht könnte man sagen, Makroevolution sei einfach eine Ansammlung von Mikroevolution. Es ist auch einfach eine Ansammlung von Chemie und Physik. Das sind unhelpful Antworten, sodass es sich vielleicht lohnt zu fragen, wie sich wissenschaftliche Domänen zueinander verhalten. Wenn ein Wissenschaftler oder Philosoph fragt, ob zwei Theorien aufeinander reduzierbar sind, gibt es mehrere Antworten. Eine ist, ob die zu reduzierende erste Theorie A durch die reduzierende Theorie B adäquat erfasst wird. Eine andere ist, dass A nicht vollständig durch B erfasst wird. Eine dritte ist, dass A und B jeweils überlappende Bereiche haben, die nur sie erfassen. Dies wird als das Problem der Theorienreduktion bezeichnet.
Reduktion ist seit etwa 60 Jahren ein philosophisches Problem im Hinblick auf die Wissenschaft. Sie tritt in drei Hauptformen auf: methodologische Reduktion, die die Auffassung ist, dass man Erklärungen für Ganzes in Bezug auf die Teile und ihre Wechselwirkungen versuchen sollte; ontologische Reduktion, die die Auffassung ist, dass alle Einheiten oder Entitäten einer Theorie aus Einheiten oder Entitäten einer anderen bekommen (zusammengesetzt sind); und metaphysische Reduktion, die die Behauptung ist, dass nur eine Art von Ding existiert (auch als "Monismus" bezeichnet). Ontologische Reduktion umfasst die Reduktion aller Gesetze und dynamischen Verallgemeinerungen der A-Theorie auf Gesetze und dynamische Verallgemeinerungen der B-Theorie. In der Wissenschaftsphilosophie wird der Fall oft genau in diesen Begriffen dargestellt, aber zunehmend beschäftigen sich Philosophen auch mit den Objekten wissenschaftlicher Theorien sowie den Modellen.
Betrachten wir Atome als Beispiel. Zu der Zeit, als Dalton Atome vorschlug, versuchte er, größere Dinge in Bezug auf kleinere Dinge zu erklären, deren Eigenschaften sich zu den Eigenschaften des Ganzen addieren. Er tat dies, weil er es für eine gute Regel hielt, das Ganze in Bezug auf die Teile zu erklären. Daher war er ein methodologischer Reduktionist, der Dinge in Bezug auf ontologische Reduktion erklärte. Er war jedoch kein metaphysischer Reduktionist, wenn er annahm, dass die Realität aus mehr als nur Atomen besteht – wie zum Beispiel Schwerkraft oder Licht (oder Gott). Ein paralleler Fall ist das genetische Reduktionismus, bei dem Verhaltensweisen auf Gene „reduziert" werden – es ist sowohl methodologisch als auch ontologisch reduktionistisch im Bereich von Verhalten und Biologie. Es behauptet jedoch nicht, dass alles in der Biologie genetisch ist, denn wir wissen, dass die Expression von Genen durch nicht-genetische Faktoren beeinflusst wird, wie beispielsweise die Verfügbarkeit von Nahrung während entscheidender Phasen der Entwicklung.
Die reduktive Beziehung zwischen Mikroevolution und Makroevolution ist heftig umstritten. Es gibt diejenigen, die mit Dobzhansky sagen, dass sich Makroevolution auf Mikroevolution zurückführen lässt. Wir können dies in drei Behauptungen unterteilen: innerhalb des „Universums" der Biologie könnte man sagen, dass alles Biologische am besten durch Mikroevolution erklärt wird (methodologisch), oder dass alle Entitäten und Prozesse der Makroevolution mikroevolutionär sind (meist genetisch – dies ist ontologisch), oder dass alles, was geschieht (in der Biologie), genetisch ist (metaphysisch). Im metaphysischen Fall erlangen Gene eine fast mystische Bedeutung, und kein seriöser Biologe macht diese Behauptung, obwohl Gegner einigen (insbesondere Dawkins) vorwerfen, dies zu tun.
Die beiden reduktionistischen Behauptungen, die wir nun betrachten werden, sind die methodologische und die ontologische.
Die methodologische Behauptung, dass Makroevolution (Ma) sich auf Mikroevolution (Mi) reduziert, ist die Behauptung, dass die optimale Lösung zur Erforschung der Evolution darin besteht, Modellierung und Tests mittels genetischer Techniken anzuwenden. Und dies hat sehr erfolgreich funktioniert. Allerdings war dies kein uneingeschränkter Erfolg – die Entwicklungsbiologie lässt sich nicht leicht auf die Genetik reduzieren, noch die Ökologie. Zellteilung, Spezialisierung und Signalisierung erklären die Entwicklung, und die Beziehung zwischen Genen und diesen Prozessen ist mehrdeutig – das heißt, einige Gene spielen eine Rolle in vielen Entwicklungsprozessen, und viele Gene spielen eine Rolle in fast allen Prozessen. Darüber hinaus sind bei der Entwicklung viele andere Faktoren beteiligt: epigenetische Faktoren (para-genetische Vererbung und umweltbedingte Modulation genetischer Effekte), zytologische Vererbung (Organellen, Zellmembranen, Ribosomen und Enzyme von Elterncellen und Elterngesellschaften). Daher reichen Gene allein nicht aus, um zu erklären, warum die Evolution entlang der Wege verläuft, die sie genommen hat. Eine Reaktion auf den methodologischen Reduktionismus in der Biologie bestand darin, zu behaupten, dass Gene lediglich „Buchhaltungs"-Entitäten für die evolutionäre Untersuchung sind (Gould 2002). Der methodologische Reduktionismus ist nicht ausreichend, selbst wenn sich herausstellt, dass Gene die einzigen signifikanten „Spieler" in der Evolution sind.
Es ist diese Annahme, die Antireduktionisten im Fall des ontologischen Reduktionismus herausfordern. Es gibt, so sagen sie, Entitäten und Prozesse, die die makroevolutionäre Dynamik beeinflussen, die in ihrer Natur jedoch nicht mikroevolutionär sind. Was könnten diese sein?
Nun, eine Liste, die Reduktionisten akzeptieren würden, umfasst den Klimawandel, geomorphologische Prozesse wie Gebirgsbildung, tektonische Isolation und Drift, Vulkanismus, außerirdische Einflüsse wie Bolidenimpakte, galaktisches Wackeln, Präzession der Erdachse und möglicherweise sogar lokale Sterne, die sich nähern und den Einfluss von Kometen und anderen Boliden auf die Erde in einem Zyklus mit einer durchschnittlichen Dauer von etwa 13 Millionen Jahren verändern. Der Punkt, den die Reduktionisten jedoch machen würden, ist, dass alles, was diese Dinge betroffen, mikroevolutionär ist – nur die Frequenzen von Genen in Populationen und so weiter. Sie dienen als Umwelt, in der sich Gene ihre Frequenz ändern (oder dies nicht tun, und die Art erlischt). Was der „Spieler" in der Mikroevolution ist, ist die Population, bestehend aus Organismen, Merkmalen und Genen; kurz gesagt, der Genpool. Nichts anderes ist wichtig.
Nonreduktionisten werden jedoch argumentieren, dass es in der Makroevolution emergente Prozesse und Entitäten gibt, die ontologisch nicht erfasst werden können. Es gibt mehrere Kandidaten für diese, die jeweils von Reduktionisten herausgefordert werden. Die Grundlage dafür ist eine Auffassung der Evolution als einer Reihe von inklusiven hierarchischen Ebenen, jede davon in gewisser Weise unabhängig von den unteren Ebenen.
 d d |
||
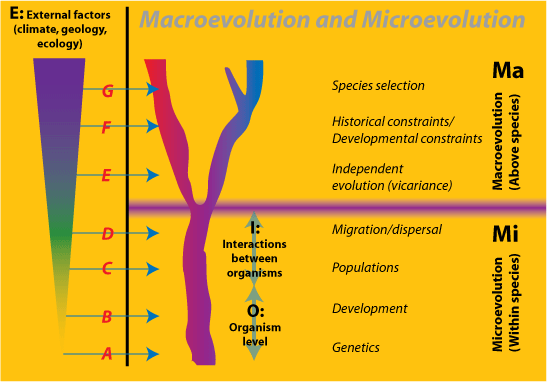
Abbildung 3. Die hierarchischen Beziehungen zwischen Makroevolution (Ma) und Mikroevolution (Mi) sowie der Umwelt (E). Mi besteht aus Organismen (O) und ihren Wechselwirkungen (I) zusammen mit Faktoren aus der Umwelt. Die Beispiele A-G illustrieren die Ebenen des Umwelteinflusses:
|
Eine Sache, die man im Hinterkopf behalten sollte, ist, dass im klassischen Reduktionismus die Richtung der kausalen Wirkung vom Mikro- zum Makrolevel verläuft. Doch Artbildung und höhere evolutionäre Prozesse beeinflussen auch, was auf niedrigeren Ebenen geschieht. David Sepkoski hat eine Tabelle, die die Art der kausalen Pfeile veranschaulicht, die sowohl nach unten als auch nach oben wirken:
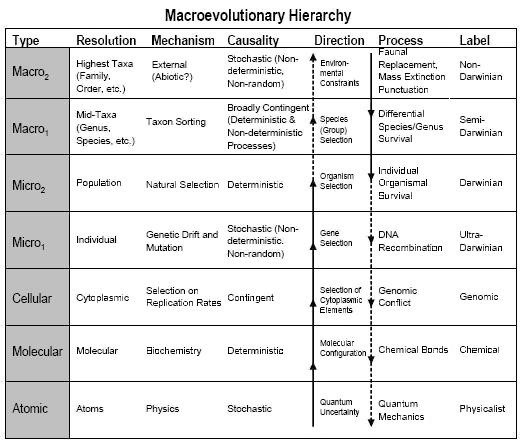
Die Häufigkeiten von Objekten wie Arten oder Organismen oder Allelen werden durch die Kontexte beeinflusst, in denen sie auftreten, welche in der Regel Prozesse auf der nächsthöheren Ebene sind. Seine Notation beschreibt klar, was in diesem Sinne als „darwinisch" bezeichnet werden kann, obwohl man nicht unbedingt akzeptieren muss, dass die Macro2-Ebene „nicht-darwinisch" ist, wenn damit etwas gemeint ist, das der Evolution im weiteren Sinne widerspricht.
Artenauswahl/-sortierung
Elisabeth Vrba (Vrba 1985, Gould und Vrba 1993) schlugen vor, dass Arten auf eine verzerrte Weise entstehen und aussterben. Generalistenarten (eurytopes) neigen dazu, länger zu überleben – wenn eine Nahrungsquelle nicht verfügbar ist, wechseln sie zu einer anderen, bis diese wieder verfügbar ist, wodurch sie Räuber-Beute-Zyklen vermeiden, die als Lotka-Volterra-Zyklen bekannt sind (wie z. B. ein drastischer Rückgang der Fuchspopulationen, wenn Kaninchen überjagt werden). Spezialistenarten (stenotopes) sind hingegen empfindlich gegenüber den kontingenten Veränderungen, die durch Klimawandel – selbst langanhaltende Dürren – erzwungen werden. Doch Spezialisten neigen dazu, häufiger zu arten, auch wenn sie häufiger aussterben, da sie sich an den Verlust oder die Verschlechterung ihrer Nahrungsressourcen anpassen. Selektion ist ein Prozess der differenziellen Überlebensfähigkeit, der mit ökologischem Erfolg korreliert; daher betrachten Befürworter dieser Art der Selektion dies als einen Selektionsprozess auf Artenebene. Andere bezeichnen es als einen Artensortierungsprozess (siehe Grantham 1995 für eine Übersicht), da Arten nicht ausreichend wie Organismen/Individuen sind. Gould veröffentlichte kurz vor seinem Tod eine ausführliche Diskussion (Gould 2002: 644-673). Es ist hier erwähnenswert, dass wenn Arten selektiert werden, dies eher der asexuellen Evolution ähnelt als der Evolution sexueller Organismen, da Arten selten durch die Rekombination von Linien entstehen, oder zumindest tierische Arten nicht. Pflanzliche Arten tun dies oft (bei etwa 5-10 % der neuen Arten), und wir haben unzureichende Beweise über andere Gruppen, um Verallgemeinerungen vorzunehmen.
Historische Einschränkungen/Baupläne
Einige "Nicht-Synthese"- oder post-Synthese-Evolutionisten glauben, dass die Prozesse, die die Artbildung verursachen, einer anderen Art angehören als diejenigen, die innerhalb von Arten stattfinden. Das heißt, sie geben zu, dass Makroevolution stattfindet, glauben aber, dass normale genetische Veränderungen durch vorgeschlagene Mechanismen wie entwicklungsbiologische Einschränkungen begrenzt werden. Diese Ansicht ist ursprünglich mit den Namen von Schmalhausen und Waddington verbunden, die von den Theoretikern der modernen Synthese oft als Nicht-Darwinianer charakterisiert wurden. Allerdings wurden mit dem jüngsten Aufschwung des als "Evo-Devo" oder evolutionäre Entwicklungsbiologie bekannten Feldes viele von Waddington und anderen vorgeschlagenen Ideen erneut untersucht (Schlichting und Pigliucci 1998, Amundson 2005, Levinton 2001).
Es gibt verschiedene Arten von Einschränkungen für die Evolution. Die bekannteste ist natürlich die selektive Einschränkung: manche Formen sind einfach nicht lebensfähig, auf welche Weise auch immer. Aber entwicklungsbedingte Einschränkungen wurden vorgeschlagen, um zu erklären, warum beispielsweise bei Tausendfüßern die Anzahl der Segmente immer eine ungerade Zahl ist (Arthur 2003). In diesen Fällen ist die Einschränkung die Natur des Entwicklungssystems selbst. Andere (Schlichting und Pigliucci 1998) betrachten dies als weitgehend ein Fall von Selektion; das Entwicklungssystem – tatsächlich die Fähigkeit zur Evolution – unterliegt ebenfalls der Selektion. Historische Einschränkungen bilden eine Art „du kommst von hier nicht dorthin"-Klasse. Sobald etwas evolviert ist, ist jeder Zustand, der eine Umkehrung der Evolution dieses Merkmals erfordert, um anderswo hinzukommen, verschwindend unwahrscheinlich. Daher werden die Dynamiken der Evolution dieses Merkmals durch das eingeschränkt, was bereits evolviert ist.
Die Vorstellung eines bauplan – eines deutschen Wortes, das "Bauplan" oder "Bauvorlage" bedeutet – wurde auf die Evolution am bekanntesten von Gould und Lewontin (1979) angewendet. Baupläne (das Wort nimmt hier den englischen Plural) sind die Körperpläne der Phyla, der zweit höchsten Linne'schen taxonomischen Ebene. Seit Georges Cuvier sie im frühen 19. Jahrhundert benannte, wurden Phyla (Singular Phylum) als distincte und natürliche Gruppierungen innerhalb der Tiere angesehen (zumindest nicht bei Pflanzen, wo die Ebene Division ist). Baupläne wurden mit der Vorstellung einer entwicklungsbiologischen und einer historischen Einschränkung in Verbindung gebracht. Es gab Kritik an der Vorstellung eines Bauplans, als mystisch in seiner kausalen Kraft. Andere sehen es als etwas, das nicht leicht durch die Prozesse der innerhalb einer Art (Mi) Evolution verändert werden kann.
Emergente Eigenschaften
Einer der von Nicht-Reduktionisten aufgestellten Behauptungen ist, dass Evolution auf emergenten Eigenschaften stattfindet. Eine emergente Eigenschaft ist eine, bei der die Eigenschaft eines Systems oder Objekts einer höheren Ebene nicht auf die Eigenschaften seiner Bestandteile reduziert werden kann, sondern stattdessen aus den Wechselwirkungen zwischen ihnen „emergiert" (O'Connor und Wong 2002, Mandik 2004). Emergente Eigenschaften wurden erstmals von einem, zufälligerweise, Freund Darwins, G. H. Lewes, im Bereich der Psychologie vorgeschlagen, aber die Idee geht auf J. S. Mill im Jahr 1843 zurück. Oft wird Emergenz als Schlagwort formuliert als „das Ganze ist mehr als die Summe seiner Teile". Emergenz wurde zum Thema, als sie ironischerweise genug in den 1920er Jahren auf Evolution angewendet wurde, von Jan Smuts und C. D. Broad.
In der Evolution wird eine Art von einigen Nicht-Reduktionisten als ein System betrachtet, das Eigenschaften aufweist, die über das Niveau des Individuums, der Verwandtschaftsgruppe oder der Deme (Zuchtpopulation) hinausgehen, basierend teilweise auf Mayrs Definition einer Art als eines geschützten Zuchtgutpools ( Mayr 1996). Dies wurde aus verschiedenen Gründen in Frage gestellt, nicht zuletzt, weil Arten in der Regel keine systematischen Wechselwirkungen zwischen allen ihren Teilen aufweisen und das angemessene Niveau die Population ist.
Nach Ansicht dieses Autors ist eine emergente Eigenschaft einfach eine Eigenschaft, die wir Schwierigkeiten haben, zu berechnen oder vorherzusagen, ausgehend von einem Wissen über die konstituierenden Teile, doch diese einfache Abweisung ist unzureichend. Wir müssen die folgenden Aspekte der Sache identifizieren:
E: Die Umweltfaktoren, in denen eine Art existiert – zum Beispiel geologische und klimatische Veränderungen
O: die Eigenschaften der Organismen – zum Beispiel ihre Merkmale und Fähigkeiten einzeln
I: die Interaktionen zwischen den Organismen – beispielsweise die Erblinien auf den Gen-, Haplotyp-, Genom- und Entwicklungsorganisationsebenen. Auch die Frage, wie Organismen ihre Umwelt durch einen Rückkopplungsprozess, der als „Nischenkonstruktion“ bekannt ist, verändern, betrifft sowohl E als auch I (Oyama et al. 2000). Wir können die reduktionistische Position wie folgt formulieren:
Reduktionismus: I(O & E) → Ma
Ma ist daher das Ergebnis der Vereinigung, auf irgendeine Weise, von E, O und I. Dies kann massiv komplex sein und „plötzliche" Veränderungen hervorrufen [Hinweis 3], oder den evolutionären Prozess über lange Zeiträume in einem Zustand der Stase halten. Ob man dies als „wiederholte Runden der Mikroevolution" bezeichnen will oder nicht (Erwin 2000), ist umstritten. Und selbst wenn es so ist, müssen wir immer noch die Modelle dafür kennen, wie sie zusammenhängen, und was Mi abdeckt.
Die alternative, nicht-reduktionistische Position besagt, dass es Eigenschaften und Prozesse gibt, die nicht allein auf E, O und I zurückgeführt werden können. Es gibt einige andere Vorgänge, nennen wir sie M, die hinzugefügt werden müssen.
Nichtreduktionismus: M & I(O & E) → Ma
Die Argumente in der Biologie befassen sich daher damit, was die Menge der Ms sein könnte und wie sie funktionieren.
Hindernisse für die Makroevolution?
Es ist eine gängige Behauptung von Antievolutionären, dass es eine Grenze für die Menge an Veränderungen gibt, die vorgenommen werden können. Kreationisten wie Gish (1979) behaupten, es gebe eine Begrenzung innerhalb der „grundlegenden Arten", ohne genau angeben zu können, was diese grundlegenden Arten sein mögen, oder warum die Veränderung innerhalb dieser begrenzt ist. Andere, wie Johnson (1991:18), behaupten, die Grenze liege in der Verfügbarkeit von genetischer Vielfalt, und dass, wenn diese Grenze erreicht ist, die Veränderung aufhört. Obwohl er zugibt, dass „Darwinisten" „einige Punkte zu machen haben", ist er kaum fair, wenn er sagt, dass Variation „möglicherweise durch Mutation erneuert werden könnte, aber ob (und wie oft) dies geschieht, ist nicht bekannt" (S. 19). Natürlich ist dies bekannt. Wir haben seit den 1910er Jahren experimentelle Belege für Mutationsraten, und moderne Forschung bestätigt sowohl mathematisch als auch empirisch, dass Mutationsraten bei etwa 0,1–1,5 pro Zygot auftreten, was bedeutet, dass jeder Embryo im Durchschnitt zwischen 1/10 und 1,5 Mutationen aufweist, abhängig von der Art (Crow 1997). Die durchschnittliche Mutationsrate – das heißt die durchschnittliche Rate persistierender Mutationen in einer Population – beträgt 2,2 x 10-9 (Kumar und Subramanian 2002). Weiterhin haben Gene keine evolutionären Geschichten, die exakt der Geschichte der Art entsprechen, in der sie existieren; ein Fachgebiet, das als Coalescence-Genetik bekannt ist, behandelt die Fähigkeit neuer Gene, über Artbildungsereignisse hinweg zu persistieren, sodass die Variabilität „verfügbar" ist, wenn sie selektiv vorteilhaft ist (Hey und Wakeley 1997). Beachten Sie, dass dies nicht bedeutet, dass Variation aufrechterhalten wird um verfügbar zu sein. Es ist einfach so, dass sie verfügbar ist, wenn sich selektive Drücke manchmal ändern.
Kreationisten behaupten oft, dass Arten nicht voneinander abstammen können, weil sich die Chromosomenzahlen unterscheiden. Menschen haben beispielsweise 46 Chromosomen, während Schimpansen 48 haben. Doch das menschliche Chromosom 2 ist das Ergebnis einer sogenannten Robertsonian-Fusion – die ursprünglichen Affen-Chromosomen 2p und 2q scheinen an ihren Enden (Telomere) verschmolzen zu sein, um das menschliche Chromosom 2 zu bilden (Williams, nicht datiert), und andere Arten, die große chromosomale Unterschiede aufweisen, können sich dennoch kreuzen (Nevo et al. 1994). DNA ordnet sich nach lokalen Sequenzen aus, nicht nach der großflächigen Chromosomenstruktur, und dies ist der Grund, warum Inversionen und Translokationen in Teilen der Sequenz weiterhin die Kreuzung ermöglichen.
Es scheint, dass es keinen einzigen Betrag an genetischer Variation gibt, der zwischen eng verwandten Arten gemeinsam ist und die Kreuzung verhindert. Bei einigen Arten reichen nur wenige Variationen aus. Bei anderen verhindert eine große Variation, wie der große chromosomale Unterschied bei Nevos Spitzmäusen, die Kreuzung nicht. Introgression, oder das Austreten von Genen über Artgrenzen hinweg, wurde bei Echsen, Pflanzen, Vögeln und Fischen beobachtet.
Zusammenfassend gibt es keine Hürde für die Entstehung von Arten. Dies mag jedoch nicht ausreichen, um zu zeigen, dass großräumige Makroevolution stattfindet, laut Autoren wie Johnson und Hitching (1982), doch die Logik hier impliziert eine kausale Kraft, die aktiv verhindert, dass Veränderungen eintreten, anstatt ein Problem mit dem Eintreten von Veränderungen. Denn wenn es genügend Veränderungen gibt, um neue Arten zu bilden, und jede Art sich leicht von ihrem Vorfahren unterscheidet, dann zeigt eine einfache Addition, dass viele Artbildungsereignisse über genügend Zeit großräumige Evolution verursachen können. Eine Reise von tausend Meilen beginnt mit einem einzigen Schritt. Umgekehrt können viele einzelne Schritte große Distanzen überbrücken. Es gibt keinerlei Hinweise auf Hürden für großräumige Veränderungen (Brauer und Brumbaugh 2001), obwohl Kreationisten gerne einige anbieten können.
Falsifizierung der Makroevolution
Ist Makroevolution eine überprüfbare Hypothese? Kann sie widerlegt werden?
Antievolutionisten versuchen darzulegen, dass Makroevolution eine Tautologie ist, genau wie sie behaupten, dass natürliche Selektion eine Tautologie sei. Die Implikation ist, dass Makroevolution nicht getestet und als falsch erwiesen werden kann und daher keine Wissenschaft ist.
Um dies zu verdeutlichen, betrachten wir, was Wissenschaftler testen, wenn sie eine Hypothese prüfen. Nehmen wir an, wir testen die Idee, dass die globale Erwärmung durch einen Anstieg von CO2 in der Atmosphäre verursacht wird. Dies besteht aus zwei Teilen – einerseits die Behauptung, dass CO2 die Speicherung von Sonnen- und anderer Wärme bewirkt, und andererseits, dass dies in der Vergangenheit eingetreten ist und tatsächlich jetzt geschieht. Wenn Sie zeigen, dass in einem bestimmten Fall keine globale Erwärmung stattgefunden hat (z. B. während der letzten Interglazialzeit), haben Sie damit nicht gezeigt, dass CO2 die globale Erwärmung nicht verursacht, noch dass es dies nicht jetzt tut. Sie haben lediglich einen bestimmten Fall getestet.
Wir können eine bestimmte Behauptung der Makroevolution testen. Wir können beispielsweise testen, ob Marder enger mit Rotpandas verwandt sind als mit Bären (Flynn und Nedbal 1998, Flynn et al. 2000). Dies ist ein Test einer bestimmten evolutionären Stammbaum oder eines Szenarios. Es testet eine historische Rekonstruktion. Wenn auf der Grundlage der Beweise und der besten Daten gezeigt wird, dass diese falsch ist, dann wurde diese Geschichte tatsächlich widerlegt. Aber können wir die Idee der gemeinsamen Abstammung testen? Es ist nicht möglich zu zeigen, dass etwas niemals vorgekommen ist, aber es ist sehr leicht zu zeigen, dass dort, wo es eintreten sollte, es entweder eingetreten ist oder nicht. Die Wissenschaft wird eine schlechte Idee nicht beibehalten, wenn wiederholt gezeigt wird, dass sie nicht erklärt, was wir berechtigt sind zu erwarten, dass sie erklärt (dies ist einer der Gründe, warum der Kreationismus in den 1850er Jahren aus der Wissenschaft entfernt wurde). Wenn Makroevolution beständig als gegen die Daten verlaufend gezeigt würde, würde die Wissenschaft sie fallen lassen und nach einer anderen Lösung suchen.
Darüber hinaus hat die Wissenschaft in gewissem Maße die ursprüngliche Vorstellung der Makroevolution widerlegt. Die ursprüngliche Idee war, dass die Evolution nur baumartige Muster bildete – Arten spalteten sich wie Äste. Eine wachsende Konsensmeinung argumentiert, dass sowohl Hybridisierung (Wiedervereinigung von Arten) als auch lateraler genetischer Transfer (Gene, die einzeln oder als Teil symbiotischer Organismen in die zelluläre Maschinerie des „Wirtstaxons" übergehen) häufiger sind, als wir zuvor dachten. Die Makroevolution von Arten wird nach wie vor als die häufigste Art betrachtet, auf der sich die Vielfalt des Lebens entwickelt hat, doch der „Baum" hat nun „Reben", die sich über die Äste einzelner Zeller erstrecken (Abb. 4).
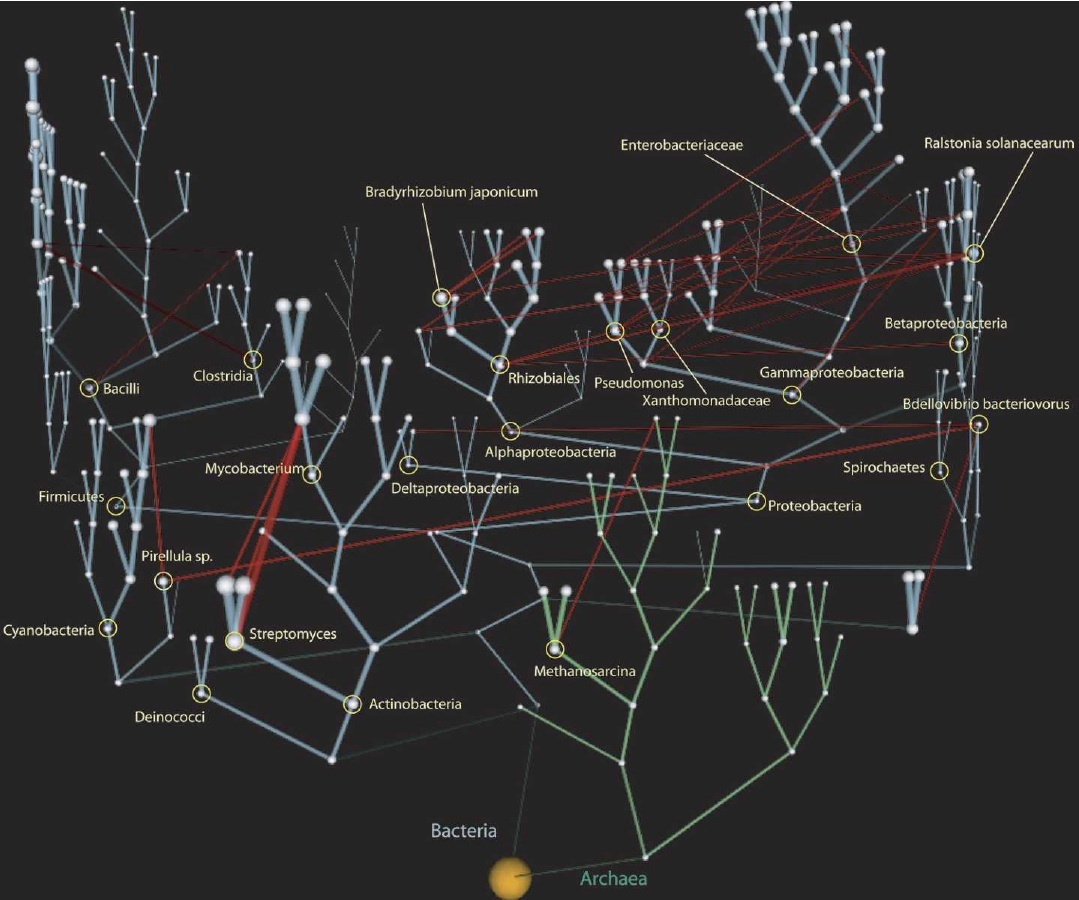 d d |
||
| Abbildung 4. Evolutionäre "Vines" Lateraler genetischer Transfer über den Baum des Lebens. Entnommen aus dem Blog von Carl Zimmer, The Loom, basierend auf Arbeiten von Victor Kunin, et al. Dieses Bild zeigt nur Bakterien und Archaeen, doch die gleichen Schlussfolgerungen gelten im größeren Maßstab anderer einzelliger Organismen. [Vollbild] |
{kind=link}
Die Hypothese der gemeinsamen Abstammung, wie wir die allgemeine Idee nennen könnten, oder die Vorstellung der Abstammung mit Modifikation, wie Darwin sie in seinem Briefwechsel nannte, wird bei jeder Prüfung einer bestimmten Hypothese getestet. Wenn es in genügend vielen Phylogenien Probleme gibt, dann kann die gemeinsame Abstammung verworfen werden. Bisher ist sie jedoch eine sehr gute erste Näherung, und die Tatsache, dass Revisionen vorgenommen werden können und auch wurden, zeigt, dass sie weder Dogma ist noch vor Daten abgeschirmt.
[Siehe 29+ Belege für Makroevolution für spezifische Falsifizierungstests.]
Fazit
Ist Mikroevolution von Makroevolution unterschieden und umgekehrt? Wir schlossen, dass dies sehr stark davon abhängt, was mit "unterschieden" und dergleichen gemeint ist. Alle Phänomene der Mikroevolution – Evolution unterhalb der Art-Ebene – müssen notwendigerweise einige Auswirkungen oberhalb der Art-Ebene haben. Ob dies ein additiver Effekt ist oder nicht, hängt von der Komplexität der Beziehungen zwischen den beiden Ebenen in jedem Einzelfall ab. Zumindest einige Makroevolution ist das Ergebnis mikroevolutionärer Prozesse. Wir fragen uns also nun, ob alle es sind. Dies ist umstritten: Die E (umweltbedingten) Faktoren, die die Makroevolution beeinflussen, sind nicht innerhalb der Art (Mi) wirkende Kräfte, sondern vermitteln mikroevolutionäre Prozesse wie Änderungen der Genfrequenzen diese notwendigerweise? Und diese Frage ist unter Spezialisten noch nicht geklärt. Eines können wir jedoch jetzt sagen: Wir können zwischen den beiden Bereichen kein einfaches Gleichheitszeichen setzen. Es ist eine offene Frage, die innerhalb der Evolutionsbiologie und verwandter Disziplinen viel diskutiert wird, ob Mi = Ma in irgendeinem Sinne gilt.
Ontologisch werden alle Objekte von Ma durch die Objekte von Mi sowie die Objekte und Prozesse von E erklärt. Wir können jedoch nicht einfach annehmen, dass alle Prozesse von Ma lediglich die Summe der Prozesse von Mi sind – dies muss nachgewiesen werden. Methodologisch können wir die Ergebnisse von Ma nicht aus dem Wissen über die Zustände von Mi plus E vorhersagen. Dies liegt nicht daran, dass die Ergebnisse von Ma nicht das Ergebnis von Mi und so weiter sind, sondern daran, dass wir diese Ergebnisse in angemessener Zeit nicht berechnen können – zu viele Variablen, Bedingungen und Verknüpfungen (Dupré 1993, Rosenberg 1994).
Aber das bedeutet nicht, dass wir sagen können, es sei unmöglich, von einer Gruppe zur anderen zu evolvieren, weil es eine Barriere gibt, wie Kreationisten behaupten. Gene und Entwicklungssequenzen sind extrem veränderbar, und bisher wurde keine Barriere gefunden, noch gibt es einen Grund, eine zu vermuten. Alle moderne Biologie akzeptiert, dass Ma durch biologische Prozesse möglich ist. Die Frage ist, auf welche Weise? Und das ist eine Angelegenheit der empirischen Untersuchung, die im Gange ist, und durch die wir neue Dinge lernen.
Die Makroevolution ist mindestens Evolution auf oder über der Ebene der Artbildung, doch es bleibt unter Wissenschaftlern eine offene Debatte, ob sie ausschließlich das Endprodukt mikroevolutionärer Prozesse ist oder ob es einen anderen Satz von Prozessen gibt, der höhere Trends und Muster verursacht. Es ist die Meinung dieses Autors, dass makroevolutionäre Prozesse lediglich die Vektorsumme mikroevolutionärer Prozesse in Verbindung mit großräumigen Veränderungen in der Geologie und der Umwelt sind, dies ist jedoch nur eine von mehreren Meinungen, die von Spezialisten vertreten wird.
Die Missbrauch der Begriffe durch Kreationisten ist ausschließlich ihre eigene Arbeit. Es liegt nicht an der Art und Weise, wie Wissenschaftler sie verwendet haben. Grundsätzlich bedeuten, wenn Kreationisten "Makroevolution" verwenden, "Evolution, der wir aus theologischen Gründen widersprechen", und wenn sie "Mikroevolution" verwenden, "Evolution, die wir entweder nicht leugnen können oder die aus theologischen Gründen akzeptabel ist".
Anmerkungen
- Das ist nicht der Fall, weil Arten sich nicht über die Zeit verändern – einige tun es. Der Grund dafür ist, dass es kein objektives und gemeinsames Kriterium dafür gibt, wann eine Art sich so sehr verändert hat, dass sie als neue Art gezählt werden kann. Die Entscheidung, eine Art als neue zu zählen, wenn es keine Aufspaltung der Art in zwei oder mehr gibt, ist eine Frage des persönlichen Geschmacks oder der Konvention, was bedeutet, dass solche Entscheidungen uns mehr über die Präferenzen des Wissenschaftlers als über die Organismen selbst aussagen.
- Die Definition dessen, was als „Art" zählt, ist selbst umstritten. Meistens wird bei sexuell fortpflanzenden Organismen das biologische Artkonzept verwendet, wonach Organismen derselben Art angehören, wenn sie sich gemeinsam fortpflanzen können und ihre Nachkommen fruchtbar sind. Dies funktioniert jedoch nicht bei asexuell fortpflanzenden Organismen und scheitert häufig bei Organismen, die über große taxonomische Lücken hinweg kreuzen können. Siehe The Index of Creation Claims CB801 für Referenzen.
- Begriffe wie „plötzlich" oder „schnell" oder „langsam" oder „abrupt" sind relative Begriffe (Loon 1999). Viel Verwirrung entsteht, wenn Menschen, die an eine Zeitskala wie die geologische gewöhnt sind, in der ein Ereignis, das 5 Millionen Jahre dauert, als „plötzlich" bezeichnet werden kann, mit Menschen sprechen, für die „plötzlich" ein paar Generationen bedeutet – was je nach Organismus ein paar Stunden, Monate, Jahre oder Jahrhunderte sein kann. Ebenso ist eine „qualitative" Veränderung relativ zu den verwendeten Messgrößen. Etwas, das auf einer Messgröße qualitativ unterschiedlich ist, kann auf einer anderen eine einfache Extrapolation sein (zum Beispiel, wenn zur Darstellung der Veränderung exponentielle Skalen verwendet werden). Ich empfehle, solche relativen Begriffe aus dieser Diskussion whenever möglich zu entfernen oder sie genau zu quantifizieren, um Missverständnisse zu vermeiden.
Referenzen Zitiert
Alexandrov, D. A. 1994. Filipchenko und Dobzhansky: Fragen der Evolutionsgenetik in den 1920er Jahren. In The Evolution of Theodosius Dobzhansky, hg. von MB Adams, Princeton: Princeton University Press. [Amazon] [Powell's] [Barnes and Noble]
Amundson, R. 2005. The changing role of the embryo in evolutionary biology: structure and synthesis, Cambridge studies in philosophy and biology. New York: Cambridge University Press. [Amazon] [Powell's] [Barnes and Noble]
Arthur, W. 2003. Entwicklungsbeschränkung und natürliche Selektion. Evolution & Development 5 (2):117-118. [PubMed]
Bowler, P. J. 1983. Der Untergang des Darwinismus: Anti-darwinistische Evolutionstheorien in den Jahrzehnten um 1900. Baltimore und London: John Hopkins University Press. [Amazon] [Powell's] [Barnes and Noble]
Brauer, Matthew J., und Daniel R. Brumbaugh. 2001. Biology remystified: The scientific claims of the new creationists. In Intelligent Design and its critics, edited by R. T. Pennock. Cambridge, MA: MIT Press. [Amazon] [Powell's] [Barnes and Noble]
Burian, R. M.: 1994. Dobzhansky über evolutionäre Dynamik: Einige Fragen zu seinem russischen Hintergrund. In The Evolution of Theodosius Dobzhansky, hg. von MB Adams, Princeton: Princeton University Press. [Amazon] [Powell's] [Barnes and Noble]
Carroll, R. L. 1997. Muster und Prozesse der Wirbeltier-Evolution, Cambridge Paleobiology Series. Cambridge, UK: Cambridge University Press. [Amazon] [Powell's] [Barnes and Noble]
Crow, James F. 1997. Die hohe spontane Mutationsrate: Ist sie ein Gesundheitsrisiko? PNAS 94 (16):8380-8386. http://www.pnas.org/cgi/content/full/94/16/8380
Dupré, J. 1993. The disorder of things: Metaphysical foundations of the disunity of science. Cambridge MA: Harvard University Press. [Amazon] [Powell's] [Barnes and Noble]
Dobzhansky, T. 1937. Genetik und der Ursprung der Arten. New York: Columbia University Press. [Amazon] [Powell's] [Barnes and Noble]
Eldredge, N. 1989. Macroevolutionäre Dynamik: Arten, Nischen und adaptive Gipfel. New York: McGraw-Hill. [Amazon] [Powell's] [Barnes and Noble]
Eldredge, N. 1995. Reinventing Darwin: The Great Evolutionary Debate. London UK: Weidenfeld and Nicholson. [Amazon] [Powell's] [Barnes and Noble]
Erwin, D. H. 2000. Die Makroevolution ist mehr als wiederholte Runden der Mikroevolution. Evol Dev 2 (2):78-84. [PubMed]
Flynn, J. J., und M. A. Nedbal. 1998. Phylogenie der Carnivora (Mammalia): Kongruenz vs. Inkonsistenz zwischen mehreren Datensätzen. Molecular Phylogenetics and Evolution 9 (3):414-426. [PubMed]
Flynn, John J., Michael A. Nedbal, Jerry W. Dragoo, und Rodney L. Honeycutt. 2000. Woher stammt der Rote Panda? . Molekulare Phylogenetik und Evolution 17 (2):190-199. http://www.msb.unm.edu/mammals/publications/Flynn2000.pdf
Gavrilets, S. 2003. Perspektive: Modelle der Artbildung: Was haben wir in 40 Jahren gelernt? Evolution Int J Org Evolution 57 (10):2197-215. [PubMed]
Gavrilets, S. 2004. Fitness landscapes and the origin of species, Monographs in population biology; v. 41. Princeton, N.J.; Oxford, England: Princeton University Press. [Amazon] [Powell's] [Barnes and Noble]
Gavrilets, S., und J. Gravner. 1997. Perkolation auf dem Fitness-Hyperwürfel und die Evolution der reproduktiven Isolation. J. Theor. Biol. 184 (1):51-64. [PubMed]
Gish, Duane T. 1979. Evolution: the fossils say no! 3. Aufl. San Diego, Calif.: Creation-Life Publishers. [Amazon] [Powell's] [Barnes and Noble]
Grantham, T. 1995. Hierarchische Ansätze zur Makroevolution – Aktuelle Arbeiten zur Artenselektion und zur Effekt-Hypothese. Annual Review of Ecology and Systematics 26:301-321. [JSTOR]
Goldschmidt, R. B. 1940 (1982). The Material Basis of Evolution, Silliman Milestones in Science. New Haven: Yale University Press. [Amazon] [Powell's] [Barnes and Noble]
Gould, S. J. 1989. Wonderful Life: The Burgess Shale and the Nature of History, New York: Norton [Amazon] [Powell's] [Barnes and Noble]
Gould, S. J. 2002. Die Struktur der evolutionären Theorie. Cambridge, Mass., Belknap Press of Harvard University Press. [Amazon] [Powell's] [Barnes and Noble]
Gould, S. J., und R. C. Lewontin. 1979. Die Spandrels von San Marco und das Pangloss-Paradigma: eine Kritik des adaptivistischen Programms. Proc R Soc Lond B 205:581-598. http://www.aaas.org/spp/dser/03_Areas/evolution/perspectives/Gould_Lewontin_1979.shtml oder http://ethomas.web.wesleyan.edu/wescourses/2004s/ees227/01/spandrels.html
Hennig, W. 1966. Phylogenetic Systematics. Übersetzt von D. D. Davis und R. Zangerl. Urbana: University of Illinois Press. [Amazon] [Powell's] [Barnes and Noble]
Hey, J., und J. Wakeley. 1997. A Coalescent Estimator of the Population Recombination Rate. Genetics 145 (3):833-846. http://www.genetics.org/cgi/reprint/145/3/833
Hitching, Francis. 1982. Der Hals des Giraffen: oder Wo Darwin irrtümlich ging. London: Pan Books. [Amazon] [Powell's] [Barnes and Noble]
Johnson, Phillip E. Darwin on Trial. 1991. Washington: Regnery Gateway. [Amazon] [Powell's] [Barnes and Noble]
Kumar, Sudhir, und Sankar Subramanian. 2002. Mutationsraten in Säugetiergenomen. PNAS 99 (2):803-808. http://www.pnas.org/cgi/content/full/99/2/803
Kunin, V., L. Goldovsky, N. Darzentas, und C. A. Ouzounis. 2005. The net of life: reconstructing the microbial phylogenetic network. Genome Res 15 (7):954-9. Epub 2005 Jun 17. http://www.genome.org/cgi/reprint/gr.3666505v1
Levinton, J. S. 2001. Genetik, Paläontologie und Makroevolution. 2. Aufl. Cambridge UK: Cambridge University Press. [Amazon] [Powell's] [Barnes and Noble]
Loon, A.J. van. 1999. Die Bedeutung von „Abruptheit" in der geologischen Vergangenheit. Earth-Science Reviews 45:209-214. [Science Direct]
Mandik, P. 2005. Emergenz. In Wörterbuch zur Philosophie des Geistes. http://philosophy.uwaterloo.ca/MindDict/emergence.html
Mayr, E. 1996. Was ist eine Art, und was ist es nicht? Philosophy of Science 2:262-277.
Nevo, E., M. G. Filippucci, und A. Beiles. 1994. Genetische Polymorphismen bei bodentüchtigen Säugetieren (Spalax ehrenbergi Superspezies) im Nahen Osten erneut untersucht: Muster und Theorie. Heredity 72 (Pt 5):465-87. [PubMed]
O'Connor, T., und H. Y. Wong. 2002. Emergente Eigenschaften. In The Stanford Encyclopedia of Philosophy, herausgegeben von E. N. Zalta. http://plato.stanford.edu/archives/sum2005/entries/properties-emergent/
Oyama, S., P. E. Griffiths, und R. D. Gray, Hg. 2000. Zyklen der Zufälligkeit: Entwicklungssysteme und Evolution. Cambridge, MA: MIT Press. [Amazon] [Powell's] [Barnes and Noble]
Raup, D. M. 1991. Extinction: bad genes or bad luck? New York: W.W. Norton. [Amazon] [Powell's] [Barnes and Noble]
Raup, D. M., und J. J. Sepkoski, Jr. 1986. Periodische Aussterben von Familien und Gattungen. Science 231:833-6. [JSTOR] [PubMed]
Rensch, B. 1959. Evolution above the species level. New York, Columbia University Press. [Amazon ]
Rice, S. H. 1995. Eine genetische Theorie der Artenselektion. J. Theor. Biol. 177 (3):237-245. http://www.webpages.ttu.edu/searice/specsel.pdf
Ridley, M. 2004. Evolution. 3. Aufl. Oxford: Blackwell Pub. [Amazon] [Powell's] [Barnes and Noble]
Rosenberg, Alexander. 1994. Instrumental biology, or, The disunity of science. Chicago: University of Chicago Press. [Amazon] [Powell's] [Barnes and Noble]
Schlichting, C. D., und M. Pigliucci. 1998. Phänotypische Evolution: Ein Reaktionsnorm-Perspektive. Sunderland, MA: Sinauer Associates. [Amazon] [Powell's] [Barnes and Noble]
Sepkoski, J. J., Jr. 1987. Umweltbedingte Trends bei Aussterben während des Paläozoikums. Science 235:64-6. [PubMed] [Science]
Simpson, George Gaylord. 1944. Tempo und Modus der Evolution. New York: Columbia University Press. [Amazon] [Powell's] [Barnes and Noble]
Smith, A. B. 1994. Systematics and the Fossil Record: Documenting Evolutionary Patterns. Oxford, OX; Cambridge, Mass., USA: Blackwell Science. [Amazon] [Powell's] [Barnes and Noble]
Stanley, S. M. 1979. Macroevolution, Pattern and Process. San Francisco: W. H. Freeman. [Amazon] [Powell's] [Barnes and Noble]
Stidd, B. M., und D. L. Wade. 1995. Ist die Artenselektion von emergenten Merkmalen abhängig? Biology and Philosophy 10:55-76. [Springer Link]
Todes, D. P. 1989. Darwin ohne Malthus: Der Kampf um das Dasein in russischer evolutionärer Denkweise, Monographien zur Geschichte und Philosophie der Biologie. New York: Oxford University Press. [Amazon] [Powell's] [Barnes and Noble]
Vrba, E. S., ed. 1985. Species und Artbildung, Transvaal Museum Monographie; Nr. 4. Pretoria, Südafrika: Transvaal Museum.
Williams, Robert. Unbestimmtes Datum. Vergleich der menschlichen und der großen Affen-Chromosomen als Beleg für gemeinsame Abstammung. Aus der Evolution Evidence Page. http://www.gate.net/~rwms/hum_ape_chrom.html
Weitere Literatur
Allmon, W. D. 1994. Taxic Evolutionary Paleoökologie und der ökologische Kontext makroevolutionärer Veränderungen. Evolutionary Ecology 8 (2):95-112. [Springer Link]
Coyne, J. A. 1992. Genetik und Artbildung. Nature 355 (6360):511-5. [PubMed]
Coyne, J. A., und H. A. Orr. 1998. Die evolutionäre Genetik der Artbildung. Philos Trans R Soc Lond B Biol Sci 353 (1366):287-305. http://pondside.uchicago.edu/ceb/Coyne&Orr_1998.pdf
Coyne, J. A., und H. A. Orr. 2004. Speciation. Sunderland, Mass.: Sinauer Associates. [Amazon] [Powell's] [Barnes and Noble]
Foote, M. 1997. Die Evolution der morphologischen Vielfalt. Annual Review of Ecology and Systematics 28:129-152. http://biology.queensu.ca/~biol440/FooteARES1997.pdf
Funk, V. A., und D. R. Brooks. 1989. Phylogenetische Systematik als Grundlage der vergleichenden Biologie. Smithsonian Contributions to Botany 0 (73):1-45. [Amazon]
Ghiselin, M. T. 1997. Metaphysics and the Origin of Species. Albany: State University of New York Press. [Amazon] [Powell's] [Barnes and Noble]
Gould, S. J. 1994. Tempo und Modus in der makroevolutionären Rekonstruktion des Darwinismus. Proc. Natl. Acad. Sci. U. S. A. 91 (15):6764-6771. http://www.pnas.org/cgi/reprint/91/15/6764
Hansen, T. F., und E. P. Martins. 1996. Übersetzung zwischen mikroevolutionären Prozessen und makroevolutionären Mustern: Die Korrelationsstruktur interspezifischer Daten. Evolution 50 (4):1404-1417. [JSTOR]
Heard, S. B. 1996. Muster im phylogenetischen Baumgleichgewicht mit variablen und evolvierenden Artbildungsrate. Evolution 50 (6):2141-2148. [abstract]
Horvath, C. D. 1997. Diskussion: Phylogenetischer Artbegriff: Pluralismus, Monismus und Geschichte. Biology and Philosophy 12 (2):225-232. [Springer Link]
Hunter, J. P. 1998. Schlüsselinnovationen und die Ökologie der Makroevolution. Trends in Evolution and Ecology 13 (1, Januar):31-36. [Springer Link]
Mooers, A. O., und D. Schluter. 1998. Anpassung makroevolutionärer Modelle an Phylogenien: Ein Beispiel unter Verwendung von Wirbeltier-Körpergrößen. Contributions to Zoology 68 (1):3-18. http://dpc.uba.uva.nl/ctz/vol68/nr01/art01
Plutynski, A. Erscheint bald. Artbildung und Makroevolution. In Anya Plutynski und Sahotra Sarkar, Hgg., Blackwell's Companion to the Philosophy of Biology.
Sepkoski, D. Erscheint bald. Makroevolution. In Michael Ruse, Hg., Oxford Handbook of the Philosophy of Biology.
Solé, R. V., S. C. Manrubia, M. Benton, S. Kauffman, und P. Bak. 1999. Kritikalität und Skalierung in der evolutionären Ökologie. Trends in Evolution and Ecology 14 (4):156-160. [PubMed]
Danksagung: Dank an Larry Moran, Matt Silberstein, Tom Scharle, Douglas Theobald, Pete Dunkelberg, Josh Zelinsky, Chris Rohrer, Erik W. und viele andere im Newsgroup für Kommentare, Kritik und offene Meinungsverschiedenheiten. Dank auch an Carl Zimmer für die Abbildung „Vines" sowie an David Sepkoski und Anya Plutynski für Exemplare ihrer bevorstehenden Artikel.