A Origem da Vida
Direitos autorais 2010
[Publicado em 31 de outubro de 2006]
Atualização 1 de julho de 2010
Resumo
Mesmo as células viventes mais simples contêm centenas de proteínas, a maioria das quais é essencial para o seu funcionamento. No entanto, tal complexidade não pode ter existido na origem da vida. Com base em pesquisas na área, propõe-se aqui como, uma vez que uma molécula genética autorreplicante existiu, a vida pode ter começado e a evolução gradual da complexidade se tornou possível, em contraste com a aparência súbita da complexidade que os criacionistas alegam ter sido necessária no início da vida. São revisadas as condições para a síntese de moléculas orgânicas na Terra primitiva e discutidos os modelos de genes-antes e metabolismo-antes. Embora a origem da homociralidade de aminoácidos e açúcares tenha sido um problema intrigante por décadas, descobertas recentes fornecem explicações plausíveis.
Outros Links:
|
Sumário
- Introdução
- Condições para a síntese de moléculas orgânicas na Terra primitiva
- Um Mundo de RNA primeiro?
- Acúmulo gradual de complexidade
- Minerais e a origem da vida
- Especificidade das reações químicas e o cenário do metabolismo primeiro
- Origem da homociralidade dos aminoácidos e açúcares
- Hiperciclos
- Perspectivas
- Notas
- Referências
A Ciência nos mostra que o universo evoluiu pela auto-organização da matéria em direção a estruturas cada vez mais complexas. Átomos, estrelas e galáxias se auto-organizaram a partir das partículas fundamentais produzidas pelo Big Bang. Nas estrelas de primeira geração, elementos mais pesados como carbono, nitrogênio e oxigênio foram formados. Estrelas de primeira geração em envelhecimento então os expulsaram para o espaço; nós, que consistimos nesses elementos, nascemos, assim, literalmente de poeira estelar. Os elementos mais pesados nasceram nas explosões de supernovas. As forças da gravidade permitiram, subsequentemente, a formação de estrelas mais novas e de planetas. Finalmente, no processo de evolução biológica de células minúsculas semelhantes a bactérias (o último ancestral comum universal, abrev. LUCA) para toda a vida na Terra, incluindo nós, humanos, formas de vida complexas surgiram a partir de formas mais simples.
Ao considerar essa autoorganização de estruturas materiais no âmbito da filosofia, pode-se concluir que isso ocorre ou porque as leis subjacentes da natureza, que precisam ser extremamente especiais para permitir isso (Rees 2001, Smolin 1999, Susskind 2006), simplesmente são como são (possivelmente no contexto de um multiverso) ou porque foram projetadas por Deus para esse propósito. Como sabemos que as leis da natureza são tão autossuficientes que, com base nelas, a complexidade de todo o universo físico evoluiu a partir de partículas fundamentais e, além disso, formas de vida complexas evoluíram de formas mais simples durante a evolução biológica, podemos razoavelmente extrapolar que elas também permitiriam que a própria vida surgisse espontaneamente, por evolução química de estruturas adequadas, independentemente de acreditarmos que essas leis foram projetadas ou não. Portanto, devemos esperar uma origem da vida por causas naturais tanto das perspectivas filosóficas teístas quanto ateístas.
O estudo experimental da origem da vida foi impulsionado pelo experimento da sopa pré-biótica de Miller (Miller 1953 , Miller-Urey experiment) que produziu aminoácidos, essenciais para a vida. Nas décadas seguintes, muita química impressionante sobre os blocos de construção da vida foi realizada, mas por muito tempo muitas questões cruciais careciam de respostas experimentais que pudessem dar esperança para direções futuras firmes. Entre estas estavam a síntese de nucleotídeos, polimerização de nucleotídeos em oligonucleotídeos, incorporação de um gene autorreplicante em células simples sobre as quais a seleção natural poderia atuar, e a origem da homociralidade de aminoácidos e açúcares. Conceitualmente, a gênese do sistema de tradução de proteínas apresentou um problema fundamental. No entanto, na última década, avanços significativos foram feitos em todas essas áreas, mesmo que os detalhes ainda sejam vagos e problemas persistam em muitas questões. As reações propostas no modelo do metabolismo primeiro, que assume o metabolismo, não os genes, na origem da vida, também se tornaram mais promissoras graças a descobertas recentes. No geral, pode-se dizer que as peças do quebra-cabeça estão começando a se encaixar de tal forma que a suposição científica de uma origem espontânea da vida a partir de matéria não-viva finalmente alcançou plausibilidade no nível da evidência experimental.
Embora a pesquisa na área apareça agora muito mais promissora do que há apenas uma década, a ciência sobre a origem da vida é, em comparação com a ciência da evolução biológica, ainda consideravelmente subdesenvolvida em seu poder explicativo. Como Richard Robinson observa (Robinson 2005): Dê a biólogos uma célula, e eles te darão o mundo. Mas além de assumir que a primeira célula deve ter surgido de alguma forma, como os biólogos explicam sua emergência do mundo pré-biótico há quatro bilhões de anos?
De fato, é uma coisa que sabemos todos os materiais de construção químicos da vida, e que o funcionamento da vida pode ser plenamente explicado pela colaboração deles em um sistema extremamente complexo. No entanto, é outra coisa inteiramente diferente como, na origem da vida, eles poderiam ter formado uma organização inicial por si mesmos, passo a passo (via quaisquer processos intermediários e blocos de construção). À primeira vista, a evolução de LUCA, um precursor das bactérias, até os humanos pode parecer brincadeira de criança em comparação: começou a partir de uma maquinaria bioquímica já tremendamente complexa e inteiramente autossuficiente e, pouco a pouco, simplesmente tornou-a ainda mais complexa.
Não se pode perder de vista que as células mais elementares que atualmente conhecemos, que não dependem permanentemente do metabolismo do hospedeiro, a bactéria Mycoplasma genitalium, possuem 482 genes codificantes de proteínas (a maioria das bactérias, como E. coli, codifica mais de 2000 proteínas diferentes), dos quais, de acordo com o estudo experimental mais minucioso até a data (Glass et al. 2006), os essenciais são 387. O estudo hipotético provavelmente mais preciso (Gil et al. 2004) coloca o número mínimo de genes em 206. Todas as proteínas produzidas a partir desses genes estão envolvidas em um emaranhado de vias de metabolismo, replicação, bem como construção e manutenção da estrutura, o que é de complexidade deslumbrante.
De fato, de outra forma, além de através de tal quantidade mínima de complexidade, como uma célula primitiva poderia ter atendido às demandas básicas mencionadas acima? Como tal rede vastamente complexa de mais de 200 proteínas poderia ter surgido por si só? Poderia-se perguntar: não teria tido que surgir de uma vez? Todavia, as evidências sugerem que toda essa complexidade pode ter evoluído, passo a passo, a partir de princípios muito simples.
2. Condições para a síntese de moléculas orgânicas na Terra primitiva
Essencial para a origem espontânea da vida foi a disponibilidade de moléculas orgânicas como blocos de construção. O famoso experimento da sopa pré-biótica de Stanley Miller (Miller 1953, experimento Miller-Urey) havia demonstrado que aminoácidos, os blocos de construção das proteínas, surgiram espontaneamente entre outras pequenas moléculas orgânicas ao reagir uma mistura de metano, hidrogênio, amônia e água em um aparelho de descarga elétrica. Essas condições foram assumidas como simulando aquelas na Terra primitiva. Já em 1922, Oparin havia proposto que a Terra primitiva possuía uma atmosfera redutora (em sua obra clássica The Origin of Life, de 1936, ele expandiu essas ideias). Observações de Júpiter e Saturno mostraram que continham amônia e metano, e inferiu-se que grandes quantidades de hidrogênio também estavam presentes lá (sabe-se agora que o hidrogênio é o principal componente atmosférico desses planetas). Essas atmosferas redutoras dos planetas gigantes foram consideradas remanescentes capturados da nebulosa solar e a atmosfera da Terra primitiva foi assumida, por analogia, como semelhante.
Sugeriu-se que apenas em uma atmosfera redutora como esta, a síntese de moléculas orgânicas, também açúcares e bases orgânicas, blocos de construção de nucleotídeos, teria sido possível em grandes quantidades (Chyba e Sagan 1992).
Pesquisas posteriores levantaram dúvidas sobre a existência de uma atmosfera redutora e sugeriram uma atmosfera neutra em vez disso; veja também (Chyba 2005), o artigo complementar a Tian et al. (veja abaixo).
No entanto, novos cálculos indicam que o hidrogênio escapou da atmosfera primitiva a uma taxa muito mais lenta do que se pensava anteriormente, resultando em uma atmosfera onde o hidrogênio era um componente majoritário (cerca de 30 %) e que, portanto, era altamente redutora (Tian et al. 2005, veja também comunicado de imprensa). Os autores mediram a produção de moléculas orgânicas através de fotólise UV nessas condições e concluem que, a uma taxa de 1010kg/ano, teria sido ordens de magnitude maior do que a taxa de síntese de compostos orgânicos em sistemas hidrotermais ou da entrega exógena de compostos orgânicos à Terra primitiva.
Outro estudo recente também apoia uma atmosfera primitiva redutora. Condritos são material primitivo da nebulosa solar e são geralmente considerados os blocos de construção da Terra e de outros planetas rochosos, asteroides e satélites. Durante e após a formação dos planetas, os gases escapam do material condrito devido à alta temperatura e pressão. Cálculos sistemáticos e detalhados sobre o que esses gases devem ter sido mostram que são principalmente o hidrogênio altamente redutor, metano e amônia — os mesmos gases dos experimentos do tipo Miller-Urey (Schaefer e Fegley 2007, veja também comunicado de imprensa). A composição dos gases variou com a temperatura apenas em grau moderado e foi encontrada como sendo em grande parte independente da pressão real sob a qual a desgasificação pode ter ocorrido, o que parece apoiar a robustez das conclusões.
Os autores mencionam que
foi descoberto que uma atmosfera redutora de metano e amônia é extremamente
vulnerável à destruição pela luz UV (Kuhn e
Atreya 1979, Kasting et al. 1983). Eles também apontam, no entanto,
que desenvolvimentos recentes sugerem que uma
atmosfera redutora é mais estável do que anteriormente acreditado:
1) Foi descoberto que a fuga de hidrogênio da atmosfera da Terra era menos
eficiente do que se pensava (referindo-se ao estudo por Tian et al. acima).
2) Observações da atmosfera de Titã, lua de Saturno, que é composta
principalmente de metano e nitrogênio, mostram que aerossóis de hidrocarbonetos
produzidos fotoquimicamente formam uma camada de névoa na atmosfera superior que protege
a atmosfera inferior da destruição fotoquímica. Tal camada de névoa também poderia
ter sido produzida na Terra primitiva a partir de metano e amônia liberados por desgasificação
(Zahnle 1986, Sagan e Chyba
1997, Pavlov et al. 2000).
No entanto, mesmo se a atmosfera da Terra primitiva teria sido neutra em vez de redutora, novos dados sugerem que a síntese eficiente de aminoácidos teria sido possível também nessas circunstâncias. O grupo de Jeffrey Bada demonstrou que, contrariamente a relatórios anteriores, quantidades significativas de aminoácidos são produzidas a partir de misturas gasosas neutras sob condições adequadas (Cleaves et al. 2008). O tamponamento com carbonato de cálcio (um material rochoso comum) impediu a redução do pH causada pelo nitrito e nitrato produzidos no aparelho de descarga elétrica utilizado, aumentando dramaticamente o rendimento. Além disso, a adição de inibidores de oxidação impediu a degradação oxidativa de aminoácidos por nitrito e nitrato, aumentando ainda mais o rendimento. Inibidores de oxidação como íons ferroso podem ter sido em excesso em relação ao nitrito/nitrato na Terra primitiva (Walker e Brimblecombe 1985). O grupo de Bada também reanalisou amostras dos experimentos de descarga elétrica de Miller dos anos 1950, simulando erupções vulcânicas ricas em vapor de água. Tais erupções teriam liberado gases redutores. Nessas amostras, os aminoácidos eram mais variados do que no clássico experimento de Miller, e os rendimentos eram comparáveis ou até mesmo superiores, indicando que mesmo se a atmosfera da Terra tivesse sido neutra, a síntese pré-biótica localizada poderia ter sido eficaz (Johnson et al. 2008, veja também artigo de notícias).
É claro que, se a vida surgiu em fontes hidrotermais de águas profundas (veja abaixo), a composição da atmosfera primitiva da Terra tornaria-se em grande parte irrelevante. Até certo ponto, isso também se aplica aos blocos de construção orgânicos entregues à Terra por partículas de poeira interestelar e em meteoritos carbonáceos.
Em uma atmosfera redutora, tornada provável com as descobertas discutidas acima, a concentração de compostos orgânicos no oceano pré-biótico pode ter sido relativamente alta (De Duve e Miller 1991, e referências therein). Além disso, localmente, uma sopa pré-biótica pode ter sido altamente concentrada por processos tão simples como, por exemplo, evaporação em poças ou lagos rasos, possivelmente com ciclos de umidade/secura de longo prazo. Deve-se ter em mente, ao avaliar todos os cenários químicos, que, devido à sua natureza, a origem da vida deve ter sido um evento muito local; isso também é importante para a questão da origem da homociralidade de aminoácidos e açúcares, veja abaixo.
Agora é amplamente aceito que, na origem da vida, não existia o atual sistema de DNA/(RNA)/proteína para a informação genética, por um lado, e catálise, regulação e função estrutural, por outro. Isso levantaria a questão: o que veio primeiro, a proteína ou o DNA? A catálise proteica sem informação genética, que permite que ela seja mantida e propagada, não é suficiente a longo prazo, e a informação genética do DNA sem catálise, necessária para a função da vida, também seria inútil.
Em vez disso, assume-se que o RNA atuou como precursor tanto das proteínas quanto do DNA, no sentido de que pode servir tanto como catalisador (como as enzimas proteicas) quanto como transportador de informação genética. Mesmo na célula moderna, os ribozimas (RNAs catalíticos) ainda desempenham um papel vital, embora limitado. No ribossomo, a síntese das cadeias peptídicas das proteínas a partir do código de RNA é realizada por ribozimas. Eles também catalisam o splicing do RNA.
A hipótese de que um chamado Mundo do RNA esteve envolvido nas primeiras etapas evolutivas da vida é agora uma visão quase universalmente aceita (Joyce 2002, Orgel 2004, The RNA World 2006). Poderia este Mundo do RNA ter estado na origem última da vida? Esta é atualmente ainda uma questão em aberto. O sistema do RNA pode ser demasiado complexo para ter surgido sem síntese por um precursor genético ou metabolismo prévio sem enzimas (opções discutidas abaixo). No entanto, embora ainda existam problemas substanciais, há agora boas pistas para processos simples e espontâneos na Terra primitiva tanto para a síntese de nucleotídeos como para a sua concatenação em oligonucleotídeos.
Por muito tempo, a síntese de monômeros de RNA sob condições pré-bióticas parecia ser um problema fundamental, uma vez que a condensação de açúcar (ribose) e base nitrogenada (purinas e pirimidinas) não funciona (Orgel, 2004). A síntese pré-biótica de purinas ribonucleotídeos ainda é incerta, mas recentemente um avanço foi feito em relação à síntese de monômeros de pirimidina ribonucleotídeos (que incorporam citosina e uracila). Agora parece, em princípio, resolvido, de uma maneira completamente inesperada. O estudo do grupo de John Sutherland (Powner et al. 2009) mostra como a natureza poderia ter montado espontaneamente monômeros de pirimidina ribonucleotídeos a partir de moléculas plausíveis pré-bioticamente, através de intermediários que contribuem átomos tanto para a porção de açúcar quanto para a porção de base dos ribonucleotídeos, evitando assim completamente um passo de condensação de açúcar e base (Fig. 1). Veja também Nature News para o impacto dessas descobertas. Embora um bom caminho para a síntese de purinas ribonucleotídeos (incorporando adenina e guanina) ainda precise ser encontrado, Jack Szostak argumenta em um comentário que acompanha o artigo (Szostak 2009) que é precisamente porque este trabalho abre tantas novas direções para a pesquisa que ele se manterá por anos como um dos grandes avanços na química pré-biótica.
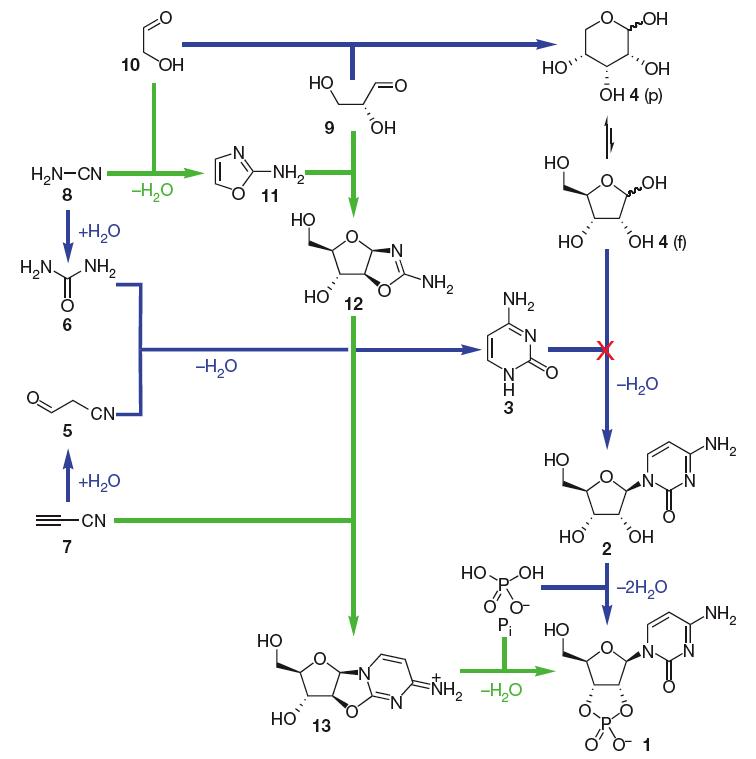
Figura 1. Montagem de pirimidina ribonucleotídica. Síntese anteriormente assumida de β-ribocitidina-2,3-cíclico fosfato 1 (azul; note a falha do passo em que citosina 3 e ribose 4 são propostas para se condensar juntas) e a nova síntese bem-sucedida descrita aqui (verde). p, piranosídeo; f, furanosídeo. De (Powner et al. 2009). Reproduzido com permissão da Macmillan Publishers Ltd.
(5 = cianacetaldeído, 6 = ureia 6, cianacetileno 7, 8 = cianamida, 9 = gliceraldeído, 10 = glicolaldeído, 11 = 2-amino-oxazol, 12 = pentose amino-oxazolina, derivado de arabinose, 13 = anidroarabinonucleosídeo)
Quanto à concatenação de nucleotídeos em oligonucleotídeos, há também progresso. A polimerização de monômeros de RNA quimicamente ativados pode ocorrer nas superfícies minerais da argila montmorilonita, gerando cadeias poliméricas de até 50-mers (Huang e Ferris 2006). Os monômeros de ribonucleotídeos pirimidínicos da nova síntese (Powner et al. 2009) também são ativados (contêm fosfato cíclico), o que pode permitir uma polimerização semelhante.
O grupo de David Deamer demonstrou que a síntese de polímeros semelhantes ao RNA pode ocorrer mesmo a partir de mononucleotídeos não ativados dentro de vesículas de fosfolipídios, devido ao potencial químico de condições anidras e hidratadas flutuantes, com o calor fornecendo energia de ativação durante a rehidratação (Rajamani et al. 2008). Tais condições poderiam ter existido ao redor de fontes termais na Terra pré-biótica. Os lipídios também fornecem um microambiente estruturalmente organizador que impõe ordem aos mononucleotídeos. Neste arranjo experimental, oligômeros de até 100 nucleotídeos podem ser formados não enzimaticamente. Restará para ver se vesículas de ácidos graxos plausíveis pré-bioticamente poderiam ter o mesmo efeito na síntese de RNA (com isso uma molécula de RNA auto-replicante também teria sido pré-empacotada para a evolução futura, cf. abaixo). A polimerização efetiva de monômeros que são ativados pode ser auxiliada por um microambiente estruturalmente organizador dentro de vesículas bem.
No entanto, embora essas reações façam polímeros semelhantes ao RNA, elas ainda não resolvem o problema da concatenação estereoespecífica 3-5 de monômeros (Orgel 2004), encontrada em todos os organismos vivos. Tanto a síntese auxiliada por lipídios quanto a polimerização em montmorilonita produzem misturas de ligações 2-5 e 3-5. No entanto, no último caso, há uma preferência pelas ligações 3-5 (até 74 %) o que é promissor. Os autores do estudo (Huang e Ferris 2006) argumentam que uma proporção maior de ligações 3-5 poderia ter resultado em uma evolução mais rápida para todas as ligações fosfodiéster 3-5 durante a replicação de oligômeros, sugerindo que talvez a especificidade completa de ligações 3-5 dentro da primeira molécula de RNA replicante não tenha sido necessária. Este cenário pode não ser impossível, dado que o RNA geralmente forma estruturas secundárias mais irregulares do que o DNA com sua hélice dupla, para a qual a estereoespecificidade absoluta da síntese é mandatória. No estudo, diferentes tipos de ativação química de monômeros de RNA resultaram em diferentes preferências para concatenação 3-5. Resta ver quão específica será a concatenação de monômeros para ligações 3-5 a partir da ativação com fosfato cíclico, o tipo de ativação relatado na nova síntese de monômeros de ribonucleotídeos pirimidínicos (Powner et al. 2009).
Devido às dificuldades da síntese de nucleotídeos de RNA e oligonucleotídeos, que, no entanto, parecem agora consideravelmente menos severas do que há apenas alguns anos, dadas as novas descobertas, várias alternativas ao RNA como o primeiro sistema genético foram propostas, que podem ter precedido-o. Ácido nucleico peptídico (PNA), ácido nucleico de treose (TNA) e peptídeos nos quais aminoácidos D- e L alternam que incorporam bases de ácido nucleico padrão (ANAs) são discutidos em (Orgel 2004). Um sistema ainda mais simples é o ácido nucleico de glicerol (GNA, veja Zhang et al. 2005). Uma ideia altamente interessante e quimicamente atraente, mas ainda não testada, é o Mundo PAH (PAH = hidrocarbonetos aromáticos policíclicos), desenvolvido por S. Platts e descrito em (Hazen 2005) e também em Wikipedia e em pahworld.com.
Outras dificuldades são que um ribozima (RNA catalítico) que possa copiar-se completamente ainda não foi encontrado até agora, um ribozima de 200 bases pode copiar cerca de 20 bases de sua sequência com alta fidelidade (Zaher e Unrau 2007), uma melhoria em (Johnston et al. 2001). Uma solução para o problema de copiar uma longa sequência de ribozima foi proposta em Szostak et al. (2001); experimentos terão que mostrar se é viável. Além disso, o formidável problema de separar o produto de fita dupla da reação de cópia de modo a permitir uma segunda rodada de cópia ainda precisará ser resolvido (Orgel 2004). Recentes descobertas sugerem que este problema pode ser abordado por ciclos térmicos, análogo à reação em cadeia da polimerase (PCR), que pode ter sido uma possibilidade sob condições pré-bióticas plausíveis (Mansy e Szostak 2008, Baaske et al. 2007).
4. Acúmulo gradual de complexidade
Vamos assumir o cenário plausível de que o RNA foi sintetizado diretamente, ver acima, de modo que, a partir de um grande reservatório de RNAs aleatórios, uma molécula de RNA autorruplicante pudesse surgir, ou que tal síntese foi realizada por um sistema genético/catalítico precursor (possivelmente na superfície de minerais, cf. Orgel 2004). Como ácidos graxos poderiam ter estado disponíveis no ambiente (Hanczyc et al. 2003, Orgel 2004), uma membrana primitiva de ácido graxo poderia ter envolvido as primeiras moléculas de RNA autorruplicantes (devido às suas propriedades moleculares, os ácidos graxos podem formar vesículas espontaneamente); isso não teria permitido a passagem dos polímeros de RNA, de modo que eles permaneceriam juntos, mas teria permitido a passagem dos nucleotídeos muito menores, fornecidos pela síntese pré-biótica espontânea ou por um sistema genético/catalítico precursor. Tal membrana teria tido características diferentes de semipermeabilidade em relação às membranas lipídicas modernas, onde a transferência de muitas moléculas é regulada através de canais proteicos.
O grupo de Jack Szostak realizou estudos extensos e plausíveis de que essas vesículas de ácidos graxos como recipientes para RNA teriam permitido crescimento e replicação meramente por mecanismos físico-químicos, até que uma maquinaria de membrana mais sofisticada, dirigida pela própria célula e mais semelhante ao que é encontrado em organismos atuais, teria tomado o seu lugar (Hanczyc et al. 2003, Chen et al. 2004, Hanczyc e Szostak 2004, Zhu e Szostak 2009).
Embora em estudos anteriores (Hanczyc et al. 2003, Hanczyc e Szostak 2004) condições mais extremas e forças brutas fossem necessárias para a divisão de vesículas, levando também à perda de uma fração substancial do conteúdo das vesículas, um novo estudo (Zhu e Szostak 2009) mostra uma solução para esses problemas. Ele utiliza vesículas multilamelares (vesículas com várias camadas de membrana lipídica) que se formam espontaneamente pela rehidratação de filmes de ácidos graxos ou pela acidificação de uma solução concentrada de micelas de ácido graxo. Uma vez formadas as vesículas multilamelares, a incorporação espontânea adicional de micelas de ácido graxo nelas causa, por meio de um mecanismo inesperado, a formação de vesículas fortemente alongadas, em forma de fio. Após a submissão a forças de cisalhamento suaves, estas dividem-se em várias, novamente redondas, pequenas vesículas filhas que preservam bem o conteúdo de RNA.
O grupo de Szostak também demonstrou que nucleotídeos podem atravessar vesículas baseadas em ácidos graxos plausíveis em condições pré-bióticas e que a cópia de modelo de uma matriz oligo dC de DNA não enzimática pode ocorrer nelas (Mansy et al. 2008), o que, em conexão com os estudos sobre o crescimento e divisão de vesículas, revela em princípio como uma protocélula heterotrófica pode ter funcionado (Fig. 2). Além disso, eles mostraram (Mansy e Szostak 2008) que membranas de modelo plausíveis em condições pré-bióticas são surpreendentemente termoestáveis, permitindo que elas tolerem pelo menos períodos curtos de temperaturas de até 100C. O ciclo térmico poderia ter sido possível perto ou dentro da superfície de chaminés hidrotermais ou fontes termais (para convecção térmica dentro de poros hidrotermais, veja também o estudo mencionado abaixo por Baaske et al. 2007). Isso poderia resolver a questão espinhosa de separar o produto de fita dupla da reação de cópia para replicação adicional. O ciclo térmico poderia ter permitido essa separação de produtos de cópia (e aumento na absorção de nucleotídeos, veja Mansy e Szostak 2008) em altas temperaturas, e cópia em temperaturas mais baixas, análogo à reação em cadeia da polimerase (PCR).
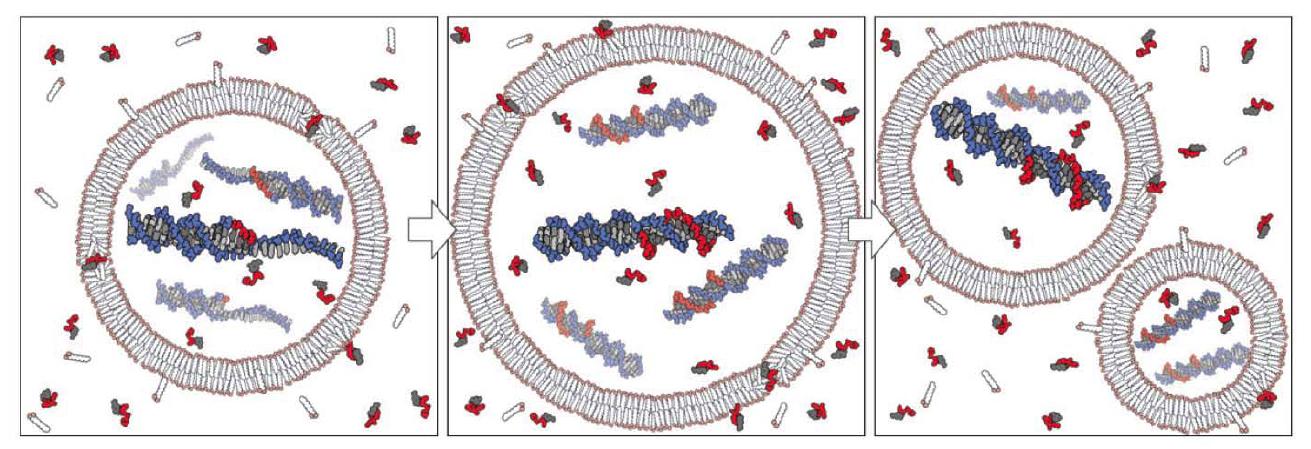
Figura 2. Modelo conceitual de uma protocélula heterotrófica. O crescimento da membrana da protocélula resulta da incorporação de anfífilos fornecidos pelo ambiente, enquanto a divisão pode ser impulsionada por forças físicas intrínsecas ou extrínsecas. Nucleotídeos ativados fornecidos externamente permeiam através da membrana da protocélula e atuam como substratos para a cópia de moldes internos. A replicação completa do molde seguida da segregação aleatória do material genético replicado leva à formação de protocélulas filhas. De (Mansy et al. 2008). Reproduzido com permissão da Macmillan Publishers Ltd.
Extrapolação A partir de todos os dados acima, dentro de vesículas de ácidos graxos, a primeira molécula de RNA autorreplicante poderia ter começado a copiar a si mesma. Durante a cópia, várias coisas teriam sido possíveis. Cópias de alta fidelidade teriam produzido a mesma molécula autorreplicante. Cópias com erros teriam resultado principalmente em RNA não funcional, mas, em uma minoria de casos, poderiam ter produzido RNA que se copiava mais rapidamente. Demonstrou-se (Chen et al. 2004, veja também artigo de notícias) que sistemas de RNA/vesículas que contêm mais material genético (o que teria resultado de uma replicação de RNA mais rápida) desenvolvem mais tensão interna do que vesículas vizinhas que não contêm tanto RNA e retiram material de membrana delas. Importante, isso teria permitido a seleção natural de vesículas por competição mesmo na ausência da capacidade de sintetizar seus próprios componentes de membrana e, portanto, de controlar diretamente seu próprio crescimento. Assim, pela primeira vez, um sistema teria tido a capacidade de sofrer evolução darwiniana pela seleção natural atuando sobre a variação. Isso teria sido uma nova e crucial propriedade emergente surgindo na transição da não vida à vida.
Uma pequena parte dos outros erros de cópia (novamente, a maioria dos erros substanciais provavelmente teria resultado em moléculas não funcionais, mas essas teriam sido filtradas pela seleção natural) poderia ter levado a moléculas de RNA com outras propriedades catalíticas inteiramente diferentes da função de cópia. Uma nova propriedade poderia ter permitido ao sistema RNA/vesícula competir ainda melhor por recursos: assim como no caso das moléculas de RNA com a melhor função de cópia, o RNA teria evoluído (1). A função de cópia das moléculas parentais provavelmente teria atuado nessas moléculas filhas também (como uma enzima RNA polimerase que copia qualquer RNA). Sistemas RNA/vesícula que possuíam as moléculas de RNA alteradas com a nova função benéfica, além de reterem o RNA com a função de cópia, teriam sido favorecidos pela seleção natural. Finalmente, através da reiteração de tais processos, uma série de novas propriedades catalíticas poderia, por exemplo, ter permitido que o pool de RNA dentro das vesículas começasse a produzir seus próprios nucleotídeos. Seria então autossuficiente? Até certo ponto, sim. Poderia isso ter sido a primeira célula primitiva? Por que não?
É apenas que, neste cenário, o metabolismo inicial teria sido muito mais simples do que o metabolismo atual: entre outras coisas, o metabolismo energético poderia ter sido substituído pela passagem de blocos de construção ativados para moléculas do ambiente externo para dentro da vesícula (num sentido, fornecendo uma substituta preliminar para a produção moderna de ATP, uma possibilidade dada o metabolismo simples), e o metabolismo lipídico, a construção da estrutura da membrana e sua regulação durante a replicação, teriam sido substituídos por vesículas simples que obedecem claramente às forças físico-químicas.
Em outras palavras: a célula teria dependido mais do mundo exterior, mas, pelo que fazia, era em certa medida autossuficiente (os organismos atuais também precisam, é claro, de nutrientes do mundo exterior). Por outro lado, a dependência da célula do mundo exterior também teria sido possível de uma maneira mais imediata. Uma célula moderna não pode, por exemplo, usar ácidos graxos do exterior de uma forma que eles sejam incorporados diretamente como elementos da membrana.
Anteriormente, perguntamos: Como uma rede complexa de mais de 200 proteínas essenciais, como é encontrada nas células mais elementares de hoje, poderia ter surgido por si só?
A chave para responder a esta pergunta parece ser a combinação das duas atribuições acima na célula primitiva: uma dependência mais direta do mundo exterior do que a célula moderna, mas também uma maior capacidade de demonstrar tal dependência ao aceitar blocos de construção moleculares como tais, sem precisar converter nutrientes neles. Partindo dessas características e gradualmente avançando a partir daí, a evolução realmente poderia ter facilitado o sistema celular a uma maior complexidade.
Talvez a síntese lipídica, em uma forma precursora da síntese moderna, tenha tornado o sistema mais independente. O sistema de RNA poderia ter, pouco a pouco, inventado a síntese proteica, como mencionado; os ribossomos modernos ainda contêm ribozimas (RNA catalítico) que catalisam a formação de ligações peptídicas, que eventualmente resultam em proteínas. Em um estudo convincente (Wolf e Koonin 2007), os autores propõem um modelo passo a passo para a origem do sistema de tradução proteica, no qual cada passo confere uma vantagem distinta a um conjunto de elementos genéticos em coevolução. O objetivo do desenvolvimento da tradução não teria sido necessário, uma visão de futuro que a evolução não possui. A causa inicial para o surgimento da tradução teria sido a capacidade de aminoácidos e peptídeos de estimular reações catalisadas por ribozimas (para peptídeos demonstrados experimentalmente, veja Robertson et al. 2004). Mesmo que se revele que vários passos na evolução da tradução provavelmente foram diferentes do modelo proposto, o estudo demonstra claramente que não há nada no surgimento do sistema de tradução que represente um caso de complexidade irredutível, incapaz de ser sujeito à evolução darwiniana passo a passo.
Um estudo recente (Bokov e Steinberg 2009), baseado na análise da interdependência dos elementos da macromolécula, mostra uma evolução passo a passo do ribossomo no nível estrutural, começando a partir de um núcleo muito pequeno, o centro de transpeptidação peptídica. Camadas adicionais foram adicionadas de fora, uma por uma, até que a atual macromolécula complexa emergisse. Cada uma dessas camadas está estruturalmente conectada apenas à respectiva anterior e poderia ser removida no modelo sem destruir a integridade das camadas internas (como a descascagem de uma cebola), demonstrando que não havia necessidade de toda a molécula emergir de uma vez. Isso estabelece a falta de complexidade irredutível da maquinaria translacional de outra perspectiva.
Finalmente, o metabolismo complexo poderia ter sido alcançado e a transição para o mundo moderno do DNA/(RNA)/proteína. O dualismo DNA/proteína, é claro, é uma fonte de complexidade em si, algo que falta em um organismo baseado apenas em RNA.
E quanto à difícil questão de um genoma que reúne todos os genes juntos? Poderia ter sido que, nas primeiras células primitivas, os RNAs foram ligados por acidente, passo a passo, um por um, formando um precursor de genoma e que cada tal passo conferisse uma vantagem na seleção natural sobre células competidoras, já que os genes não teriam sido perdidos mais durante a divisão celular e a replicação teria sido sincronizada. Com o tempo, um pequeno genoma de RNA inteiro poderia ter se organizado dessa maneira, até que mecanismos para expansão interna, como os encontrados em genomas modernos, pudessem assumir o controle, por exemplo, duplicação gênica e variação do gene duplicado.
Certamente, a leitura de uma sequência de RNA ligado como vários genes únicos teria exigido que o mecanismo celular primitivo tivesse encontrado uma maneira de reconhecer o início e o fim de uma sequência. Regiões promotoras e códon de início/parada, na forma em que são usados em genes codificadores de proteínas, não teriam estado presentes em um organismo de RNA primitivo.
Embora a falta de fidelidade de replicação teria sido um problema em genomas de RNA primordiais, a análise de estudos experimentais de mutação em ribozimas indica que um genoma de RNA poderia, no entanto, ter crescido até o tamanho de 100 genes (Kun et al. 2005, Poole 2006). O genoma de RNA poderia, pouco a pouco, ter sido substituído por um genoma de DNA, uma vantagem selecionável que as células primordiais teriam encontrado por acaso.
Quanto ao código genético universal: há evidências de que, até o último ancestral comum, o código genético foi selecionado, em grande medida, para minimizar erros, sendo, portanto, não arbitrário. No entanto, em parte, provavelmente também é um acidente congelado, uma vez que a seleção para essa característica não parece ser máxima (Koonin e Novozhilov 2009).
Em resumo, com base nos dados disponíveis, uma origem espontânea da vida como células simples contendo um único polímero genético, sobre o qual a seleção natural poderia atuar, é viável. Uma transição evolutiva gradual dessas células para uma complexidade celular comum teria sido possível.
5. Minerais e a origem da vida
Os minerais provavelmente fornecem a pista para uma série de questões relacionadas à origem da vida. Como discutido, demonstrou-se que eles catalisam a polimerização de moléculas semelhantes a nucleotídeos (Huang e Ferris 2006, Orgel 2004). Também foi demonstrado que eles permitem a síntese pré-biótica, em pureza suficiente, de um componente de nucleotídeo que se mostrou elusivo: minerais de borato estabilizam a ribose (Ricardo et al. 2004, veja também artigo de notícias; no entanto, para uma síntese de monômeros de ribonucleotídeos sem intermediário de açúcar, veja acima). A formação de vesículas de ácidos graxos também é auxiliada por minerais, e partículas minerais poderiam ter acabado dentro das vesículas e ali exibido propriedades catalíticas (Hanczyc et al. 2003, Hanczyc et al. 2007).
Em um cenário hipotético diferente, os minerais também desempenham um papel interessante. Em vez de em uma sopa pré-biótica aquosa na ou perto da superfície da Terra, tem sido hipotetizado que a vida pode ter começado nas profundezas do oceano, no ambiente único das fontes hidrotermais de águas profundas. Demonstrou-se que moléculas orgânicas relevantes podem ser sintetizadas em reações envolvendo gases e minerais presentes nesses locais, com CO e outras pequenas moléculas contendo carbono como fonte de carbono. Essas reações requerem altas temperaturas, ou uma combinação de altas temperaturas e pressões, encontradas nas fontes hidrotermais de águas profundas. Para citar alguns exemplos: a reação de CH3SH com CO na presença de FeS/NiS resulta em ácido acético ativado, ou uma mistura de H2S, FeS/NiS e CO (como única fonte de carbono) produz ácido acético (Huber, Wchtershuser 1997). A piruvato é formada a partir de CO na presença de noniltiol e FeS (Cody et al. 2000). Ácidos alfa-hidroxí e alfa-amino são sintetizados a partir de CO, ligantes ciano e ligantes metiltio sob catálise por precipitados Fe-Ni, na presença de hidróxido de cálcio ou magnésio (Huber, Wchtershuser 2006). Para críticas às condições no último estudo, bem como respostas, consulte (Bada et al. 2007).
Além disso, em água de alta pressão e alta temperatura, como a encontrada em fontes hidrotermais de águas profundas, moléculas orgânicas, uma vez formadas, apresentam um nível de reatividade química (embora nem sempre particularmente específica) que é geralmente observado em ambientes aquosos normais apenas ao acelerar as taxas de reação por enzimas (veja, por exemplo, a revisão Hazen et al. 2002). Para as propriedades físico-químicas da água de alta pressão e alta temperatura, veja Basset M-P 2003 (a água comporta-se mais como um solvente orgânico apolar nessas condições). A catálise por minerais, como os presentes em fontes hidrotermais de águas profundas, aprimora ainda mais as reações de moléculas orgânicas em tal ambiente aquoso (2). A degradação de moléculas orgânicas sintetizadas nessas condições de alta temperatura e alta pressão pode ser evitada por minerais também, pelo menos isso já foi demonstrado para aminoácidos (veja Hazen et al. 2002). Ácidos graxos, como fonte de material para formação de membranas, também poderiam ter sido sintetizados em fontes hidrotermais (Orgel 2004).
Além disso, existem também, apenas de temperatura morna a moderadamente quente, fontes hidrotermais que possuem microambientes semipermeáveis de dimensões celulares (imitando uma membrana lipídica), que poderiam reter moléculas em altas concentrações. Isso proporcionaria uma possível solução para o problema da concentração (Russell e Martin 2004, Robinson 2005). No entanto, ainda resta ver de que maneira tais compartimentos imóveis teriam permitido a seleção natural, que é possível no caso de vesículas de ácidos graxos por competição por material de membrana (Chen et al. 2004) e outros recursos, como discutido aqui. Koonin e Martin (2005) propõem que, se os micro-compartimentos minerais tivessem uma certa porosidade, unidades evolutivas mais competitivas poderiam ter se espalhado sobre outras por ocupação preferencial de novos compartimentos vazios que deveriam se formar continuamente por precipitação de sulfeto em fontes hidrotermais. Isso teria sido um progenitor da divisão celular. A alternativa que, em contraste com este modelo interessante, mas ainda hipotético, já goza de extenso suporte experimental teria sido, naturalmente, a divisão de vesículas de ácidos graxos contendo polímeros genéticos por forças físico-químicas simples, ver acima. Os autores também argumentam (cf. Martin e Russell 2003) que micro-compartimentos minerais em vez de lipídios poderiam ter servido como membrana celular mesmo para um organismo tão avançado como o LUCA, já que, em sua visão, isso permitiria a explicação mais simples do porquê os descendentes do LUCA, arqueobactérias e eubactérias, possuem membranas lipídicas muito diferentes, as quais teriam surgido apenas mais tarde.
As fontes hidrotermais podem oferecer ainda outra solução para o problema de concentração dos componentes-chave na origem da vida. Monômeros de RNA e oligonucleotídeos podem ter sido acumulados em grande quantidade em sistemas de poros hidrotermais devido a um gradiente térmico através do poro (Baaske et al. 2007, veja também o comentário acompanhante, Koonin 2007). Até 1000 vezes a concentração de nucleotídeos é alcançada em um único poro, e mais de 108 vezes em poros concatenados (esses poros podem chegar a milímetros de tamanho, sendo muito maiores que os micro-compartimentos acima mencionados). Essa acumulação não depende de superfícies de adsorção que podem ser restritivas para sequências de reações. O grupo de Jack Szostak (Budin et al. 2009) mostrou que também ácidos graxos são concentrados em grande quantidade em sistemas semelhantes. Ao concentrar soluções diluídas de DNA e ácidos graxos, a auto-organização de grandes vesículas contendo DNA encapsulado ocorre em regiões dos capilares onde a concentração crítica de agregação do ácido graxo é excedida.
Todos esses cenários de concentração podem não funcionar apenas em fontes hidrotermais de águas profundas, mas também em sistemas hidrotermais encontrados em águas rasas na periferia de vulcões. A convecção térmica dentro dos poros hidrotermais (Baaske et al. 2007) também teria permitido o ciclo térmico para separar o produto de fita dupla das reações de cópia de RNA.
A química nesses ambientes pode abrir novas possibilidades para a síntese de um polímero genético anterior ao RNA, ou do próprio RNA, necessário no cenário gene-first.
6. Especificidade das reações químicas e o cenário do metabolismo primeiro
Alguns estendem as conclusões acima mencionadas das fontes hidrotermais de águas profundas para uma variante do cenário metabolismo-antes, em oposição ao cenário gene-antes descrito acima, uma hipótese que afirma que ciclos metabólicos complexos poderiam ter se auto-organizado, independentemente de um sistema genético capaz de fornecer catalisadores poliméricos. De acordo com esta hipótese, apenas por meio de tal metabolismo estável, a síntese de nucleotídeos e oligonucleotídeos, necessários para o início do mundo do RNA, teria sido possível; no entanto, devido às recentes descobertas aqui discutidas, a síntese pré-biótica espontânea de RNA está agora se tornando muito mais provável do que quando esses cenários de metabolismo-antes foram originalmente propostos.
Nas mesmas publicações em que Günter Wächtershäuser havia previsto a síntese de moléculas orgânicas sob condições encontradas em fontes hidrotermais, previsões que foram experimentalmente confirmadas (veja acima), ele também introduziu um modelo metabólico (Wächtershäuser 1988, Wächtershäuser 1990). Esta hipótese inventiva e detalhada integra de forma impressionante uma multitude de observações no campo da química. Sugere que o início da vida foi o que tem sido denominado vida plana, um metabolismo elaborado bidimensional nas superfícies minerais de fontes hidrotermais de águas profundas; isso também aborda o problema da diluição concentrando toda a química na superfície. Central para esta e outras hipóteses de metabolismo-antes é o ciclo do ácido cítrico redutor (ciclo de Krebs reverso), que fornece um mecanismo central de biomoléculas úteis a partir de CO2 (Morowitz et al. 2000, Smith e Morowitz 2004).
No entanto, o desenrolar deste e de cenários relacionados (por exemplo, Smith e Morowitz 2004) exigiria uma surpreendente falta de reações laterais (enquanto um nível residual pequeno de reações laterais poderia favorecer o desenvolvimento evolutivo). Não é fácil ver como a extraordinária especificidade das reações químicas, necessária para sequências complexas de reações e metabolismo para o funcionamento da vida, seria, em geral, possível sem polímeros catalíticos que apresentem uma bolsa de substrato tridimensional. Estes são fornecidos apenas por um cenário de genes primeiro. Leslie Orgel (Orgel 1998) comenta sobre a questão da seguinte maneira (para uma crítica estendida, veja Orgel 2000):
Não há consenso sobre o grau até o qual o metabolismo poderia desenvolver-se independentemente de um material genético. Na minha opinião, não há base na química conhecida para a crença de que longas sequências de reações podem se organizar espontaneamente e todos os motivos para acreditar que não podem. O problema de alcançar especificidade suficiente, seja em solução aquosa ou na superfície de um mineral (3), é tão severo que a chance de fechar um ciclo de reações tão complexo quanto o ciclo do ácido cítrico reverso, por exemplo, é negligenciável. O mesmo, creio, é verdadeiro para ciclos mais simples envolvendo pequenas moléculas que podem ser relevantes para as origens da vida e também para ciclos baseados em peptídeos.
Assim, parece razoável assumir que o desenvolvimento de ciclos e vias metabólicas teria exigido polímeros genéticos/catalíticos, mesmo que, obviamente, as opiniões estejam divididas sobre o assunto.
Os grandes conjuntos autocatalíticos de Stuart Kauffman (Kauffman 1993) podem ser excessivamente otimistas pelo mesmo motivo. Os conjuntos autocatalíticos podem ter maiores chances de serem realizados quando são pequenos.
No entanto, desenvolvimentos mais recentes podem ser vistos como levantando a possibilidade de que o cenário do metabolismo primeiro seja, afinal, realista. Foi descoberto (Zhang e Martin 2006) que três dos cinco passos redutivos do ciclo reverso de Krebs (Fig. 3) podem ser impulsionados por partículas de ZnS (fornecendo poder redutor de elétrons da banda de condução e acreditado ter sido prevalente nas águas da Terra primitiva) sob a influência de luz UV. As conversões de oxalacetato a malato e de fumarato a succinato prosseguiram com rendimentos surpreendentes (75 % e 95 %, respectivamente), o que pode estar próximo do suficiente. A conversão de succinato a oxoglutarato também prosseguiu, embora a uma taxa baixa de 2,5 %. Os autores sugerem que um conjunto mineral mais complexo do que apenas ZnS pode impulsionar todo o conjunto de reações.

Figura 3. Ciclo de Krebs reverso. Em destaque estão as cinco reações de redução (rotuladas 1-5). De (Zhang e Martin 2006). Reproduzido com permissão. Copyright 2006 American Chemical Society.
De alto interesse estão também as descobertas recentes que foram relatadas (Robinson 2005) para dar esperança aos adeptos do modelo do metabolismo primeiro. Demonstrou-se (Cordova et al. 2005a) que tais moléculas orgânicas simples como aminoácidos únicos podem catalisar a síntese estereoespecífica de açúcares a partir de materiais de partida simples com especificidade semelhante à das enzimas, embora apenas em solventes orgânicos. Por exemplo, seja L- ou D-enantiômeros de certos aminoácidos (com a serina entre eles, veja abaixo) podem desencadear a formação de um certo tipo de açúcar não apenas com excelente quimioseletividade (i.é., evitando reações laterais indesejadas), mas também a formação de um entre 16 enantiômeros possíveis deste açúcar com aproximadamente 99% de estereoespecificidade. Estas reações baseiam-se no princípio da organocatálise assimétrica descoberto na década de 1970 (Eder et al. 1971, Hajos e Parrish 1974).
Ainda mais promissoras são as descobertas de um estudo de acompanhamento (Cordova et al. 2006). Aqui, os autores relatam que pequenos peptídeos (principalmente dipéptidos testados, ou peptídeos com no máximo cinco aminoácidos) (4) e tetrazóis de aminoácidos podem catalisar reações de aldol, alguns deles produzindo açúcares, com grande estereoespecificidade em água, não apenas em solventes orgânicos.
A catálise altamente específica por aminoácidos ou outras pequenas moléculas orgânicas (5) poderia, mais geralmente, ou seja, não apenas nesta reação particular, substituir, em certa medida, as bolsas tridimensionais de substrato dos polímeros catalíticos? Poderia fechar ciclos metabólicos?
Essas possibilidades especulativas, envolvendo catálise específica por moléculas orgânicas, aproximariam-se do cenário do metabolismo primeiro proposto por Christian De Duve (De Duve 1995). Ele assume um protometabolismo envolvendo um mundo de tioéster que também fornece energia para reações moleculares que teriam se tornado a base para o surgimento do mundo do RNA (este cenário foi sugerido para a sopa pré-biótica, não para a química em fontes hidrotermais). No entanto, De Duve baseia sua catálise em multímeros derivados de tioésteres, estruturalmente semelhantes às [primeiras] pequenas proteínas catalíticas. É difícil imaginar como tais unidades catalíticas relativamente grandes e complexas poderiam ter se formado constantemente com alta reprodutibilidade de maneira espontânea. Tal reprodutibilidade seria necessária para estabelecer e manter um metabolismo estável, ou, de outra forma, sequências complexas de reações, é claro, não constituem problema para polímeros genéticos, conforme fornecido pelo cenário do gene primeiro. Ainda assim, a reprodutibilidade (e abundância) da síntese sem genes de supostos catalisadores de pequenas moléculas provavelmente teria sido um problema muito menor.
O ambiente aquoso de alta pressão e alta temperatura dos ventos hidrotermais de águas profundas, que produz mudanças drásticas na reatividade de compostos orgânicos (veja acima), também pode fazer com que pequenas moléculas orgânicas atuem como catalisadores específicos que não desempenhariam essa função em solução aquosa normal?
Finalmente, para resolver o principal
quebra-cabeça do modelo gene-first: a catálise por pequenas moléculas orgânicas
poderia estar envolvida na síntese de nucleotídeos e oligonucleotídeos na forma
de estereoisômeros corretos – uma química muito mais complexa do que apenas a
síntese de açúcares – na ausência de polímeros genéticos/catalíticos precursoros?
Pesquisas futuras podem nos informar sobre todas essas questões.
Deve-se notar que a dependência da luz UV
para as reações descritas do ciclo cítrico redutor (Zhang e Martin 2006), possivelmente em conjunto com a necessidade de temperaturas moderadas sob as quais essas reações foram relatadas como ocorrendo, confinaria essa química à superfície da Terra. Como
Leslie Orgel aponta (Orgel 2006), embora historicamente os defensores do metabolismo-first tenham argumentado predominantemente pela síntese em fontes hidrotermais de águas profundas e os defensores do gene-first por uma sopa pré-biótica na superfície da Terra, nenhuma dessas conexões pode ser um pré-requisito a priori para o respectivo modelo. O metabolismo sem genes poderia ter se desenvolvido na superfície da Terra, ou genes sem metabolismo prévio em fontes hidrotermais.
Por outro lado, embora ambas as reações do ciclo do ácido cítrico redutor e a nova síntese de monômeros de ribonucleotídeo pirimidina ativados (Powner et al. 2009) sejam relatados como exigirem luz UV, reações aparentemente confinadas a um único local podem ser demonstradas, conforme o conhecimento avança, que também ocorrem em outros locais sob diferentes condições. A síntese de monômeros de RNA em sistemas hidrotermais poderia ser um cenário atraente, pois, como discutido acima, poros hidrotermais permitiriam uma grande acumulação de monômeros e oligômeros de RNA por gradientes térmicos através deles (Baaske et al. 2007).
7. Origem da homochiralidade de aminoácidos e açúcares
A vida sintetiza quase exclusivamente L-aminoácidos e D-açúcares. Uma questão fundamental relacionada à origem da vida é o surgimento dessa homociralidade (unidirecionalidade). A homociralidade é essencial para o funcionamento das proteínas como polímeros de aminoácidos e para a estrutura do DNA e do RNA, que requer a incorporação de D-açúcares.
Como surgiu essa homociralidade de aminoácidos e açúcares? É uma questão que tem intrigado pesquisadores da origem da vida por décadas, no entanto uma série de descobertas recentes aborda o problema de maneira surpreendentemente bem.
Enantiômeros (formas L- ou D-) de aminoácidos podem ser fortemente enriquecidos em duas etapas:
1. Um desequilíbrio inicial das formas enantioméricas de um aminoácido
Como uma maior presença de uma forma enantiomérica de um aminoácido em relação à outra (excesso enantiomérico, abreviado: ee), por menor que seja, poderia ter surgido de alguma forma? Afinal, a síntese típica de um aminoácido em laboratório resulta em uma proporção exata de 1:1 entre os enantiômeros L e D – uma mistura racêmica.
Uma possível fonte são os meteoritos. No meteorito de Murchison, um exemplo bem estudado, foi relatado que a forma L de alguns dos aminoácidos encontrados está presente em até 9 % ee (Cronin e Pizzarello 1997). Esses ee podem ter sido induzidos por luz UV polarizada circularmente (Bailey et al. 1998). Um estudo recente (Glavin e Dworkin 2009) relata ee de L-isovalina superiores a 18 % no meteorito de Murchison e postula como causa alteração aquosa (um processo no qual mudanças nos minerais ocorrem influenciadas pela água). Aminoácidos como a isovalina, que não é comum na biosfera terrestre, podem ter transferido quiralidade para compostos biogênicos como catalisadores em reações químicas (Pizzarello e Weber 2004).
No entanto, embora a evolução química da vida primitiva possa muito bem ter se baseado em material meteorítico, as fontes de excessos enantioméricos ligeiros ou mesmo pronunciados de aminoácidos poderiam ter surgido em diversos lugares da Terra pré-biótica.
Robert Hazen e colaboradores descobriram (Hazen et al. 2001) que cristais do mineral comum formador de rochas calcita (CaCO3) podem preferencialmente adsorver as formas D- ou L- do ácido aspártico (e em experimentos preliminares, as formas D- ou L- da alanina também) dependendo da quiralidade da superfície do cristal. A média de ee foi de alguns pontos percentuais. De forma semelhante, a adsorção de ácido 3-carboxi adipico, uma molécula relevante pré-bioticamente, em cristais de calcita ou feldspato produziu ees de até 10 % (Castro-Puyana et al. 2008).
Embora tais pequenos excessos enantioméricos locais possam ser suficientes para desencadear eventos que os amplifiquem fortemente (veja abaixo), outro estudo encontrou preferências muito maiores de adsorção de aminoácidos a superfícies minerais (Wedyan e Preston 2005). Enquanto em seu estudo quartzo, caulim e montmorilonita mostraram preferências leves para a adsorção de enantiômeros, sedimentos comuns de estuários exibiram forte seletividade. As razões típicas D/L eram enormemente diferentes de 1, chegando em seus extremos mais altos a 100 para a serina. Os sedimentos foram incinerados para remover matéria orgânica que poderia introduzir viés quiral. Os autores são cautelosos: A possibilidade de que o processo de incineração realmente esculpa e ative a superfície mineral conforme queima a matéria orgânica natural (quiral?) não pode ser descartada. No entanto, eles também observam que é notável que tal forte seletividade ocorra de qualquer forma.
Um mecanismo inteiramente diferente e atraente para alcançar excesso enantiomérico foi relatado (Kojo et al. 2004), que envolve a formação de uma fase sólida de cristais de aminoácidos. Isso poderia muito bem ter ocorrido em uma paisagem pré-biótica pelo resfriamento de uma solução aquosa concentrada e quente de aminoácidos ou simplesmente pela evaporação lenta de uma solução. A maioria dos aminoácidos racêmicos forma cristais que também são racêmicos. No entanto, quando a asparagina D, L-racêmica forma cristais, eles não são racêmicos, mas apresentam graus variados de excesso de qualquer um dos enantiômeros L ou D. Quando outros aminoácidos estão presentes, seja sua forma L ou sua forma D cristaliza preferencialmente com a forma enantiomérica (L ou D) de asparagina que confere um excesso durante a formação do cristal em questão.
O comportamento dos aminoácidos no estudo mostrou grande consistência. Se havia um viés em favor da L-asparagina em um cristal, praticamente todos os outros 12 aminoácidos co-cristalizados também mostravam um viés em favor da forma L. O grau de viés em favor da L-asparagina variava entre os cristais; quando o viés era mais pronunciado, o viés em favor da forma L nos outros aminoácidos também era mais pronunciado. Os efeitos observados podiam ser muito altos. Se em outro cristal havia um viés em favor da forma D de asparagina, os outros aminoácidos co-cristalizados exibiam também um viés em favor do seu enantiômero D.
É claro que o equilíbrio enantiomérico geral da mistura inteira de aminoácidos (a soma de todos os cristais e da fase líquida acima deles) permanecia racêmico. No entanto, se esses tipos de cristais fossem formados a partir de uma solução na Terra pré-biótica e alguns dos cristais fossem então fisicamente separados de outros — o que poderia ter ocorrido por muitos processos comuns — e posteriormente redissolvidos, os aminoácidos em solução automaticamente exibiriam um ee. Dependendo do cristal particular do qual a solução derivou, esse ee poderia ter sido alto.
Outra maneira de alcançar excesso enantiomérico dentro de um racemato são condições especiais de cristalização muito longe do equilíbrio. Normalmente, os ácidos aspártico e glutâmico formam cristais racêmicos. No entanto, quando são submetidos à ascensão capilar através de um material poroso (como um tijolo parcialmente imerso em uma solução), o que é acompanhado por supersaturação devido à evaporação da solução através da estrutura porosa, cristais puros L- ou D- são formados (Viedma 2001). Condições semelhantes podem ter estado presentes na Terra primitiva em ambientes sedimentares, como em uma playa ou barra de areia.
2. Aumento do excesso enantiomérico por equilíbrios de fase sólida-fase líquida
Estudos demonstraram que, uma vez que exista um excesso inicial de um enantiômero em uma mistura de aminoácidos, mesmo que seja apenas muito leve, ele pode ter um efeito enorme. Este efeito pode ocorrer quando aminoácidos sólidos e dissolvidos de tal mistura coexistem em equilíbrio, ou seja, quando cristais se formam, por exemplo, devido à evaporação limitada de uma solução.
Um estudo detalhado foi realizado pelo grupo de Donna Blackmond (Klussmann et al. 2006). Quando uma mistura de enantiômeros L- e D- de um aminoácido mostra um excesso de um enantiômero, na maioria dos casos, um equilíbrio entre o aminoácido sólido e dissolvido consistirá nos seguintes dois ou três componentes:
a) cristais racêmicos (cristais com uma proporção de 1:1 dos enantiômeros L- e D-, sem excesso enantiomérico em nenhum dos lados)
b) cristais puros do enantiômero em excesso
(Se apenas uma dessas duas fases sólidas estiver presente, ou se ambas coexistirem, dependerá do ee global.)
c) uma solução de aminoácido em equilíbrio com a(s) fase(s) sólida(s), que também exibe um certo ee.
No entanto, como se revela, para vários aminoácidos, esse ee em solução é muito maior do que o ee geral da mistura total (o material sólido, dominado por cristais racêmicos, mostra, portanto, um ee correspondentemente menor). No caso mais extremo, a serina fornece uma solução quase enantiopura (> 99 % ee) em água a partir de uma amostra quase racêmica (apenas cerca de 1 % ee) sob condições de equilíbrio sólido-líquido.
Um estudo menor, conduzido independentemente em torno do mesmo período, relata resultados semelhantes (Breslow, Levine 2006). A evaporação lenta de uma solução aquosa de fenilalanina com apenas 1 % ee do enantiômero L resultou em uma solução desse aminoácido com 40 % ee do enantiômero L acima do material sólido. Se, por sua vez, tal solução fosse deixada evaporar, a solução resultante em equilíbrio com o material sólido apresentaria 90 % ee.
Em um estudo mais recente, o grupo de Blackmond (Klussmann et al. 2007) expandiu o conceito para misturas de aminoácidos com outros compostos, que podem co-cristalizar com os aminoácidos. Eles mostraram que, ao influenciar a solubilidade, em alguns casos esses compostos influenciaram fortemente a ee em solução sob condições de equilíbrio sólido-líquido. Por exemplo, sob essas condições, a ee da valina foi aumentada de 47 % até 99 % na presença de ácido fumárico. Observe que a plausibilidade pré-biótica é aprimorada neste cenário, pois ele emprega misturas de compostos em vez de componentes puros.
Outra maneira de aumentar o ee não emprega enriquecimento de ee em solução, mas na fase sólida de um equilíbrio sólido-líquido. Um mecanismo curioso de crescimento de cristais, que envolve abrasão por moagem, permite a emergência de um único estado quiral na fase sólida a partir de um desequilíbrio inicial na composição do cristal tão baixo como 2-3 % ee (Noorduin et al. 2008). No entanto, requer racemização do composto em solução e foi aplicado a um derivado de aminoácido que sofre essa racemização após adição de base. Uma explicação teórica diferente daquela fornecida no estudo é discutida em um comentário sobre as descobertas (McBride e Tully 2008). Em um estudo de acompanhamento, as descobertas foram estendidas a um aminoácido próprio, o ácido aspártico (Viedma et al. 2008). A racemização em solução foi realizada com salicilaldeído catalítico em meio ácido, e até 99 % de ee em cristais foi obtido a partir de um desequilíbrio inicial de menos de 10 % de ee. O aquecimento poderia substituir a moagem mecânica.
Nos estudos de Klussmann et al. 2006 e de Breslow, Levine 2006, as descobertas são discutidas também em termos de catálise por aminoácidos de reações de aldol, que incluem a síntese de açúcares e podem resultar em D-açúcares enantiopuros; veja também as descobertas de Cordova et al. 2005a e Cordova et al. 2006 revisadas acima. No entanto, quando se considera a nova via de síntese para monômeros de ribonucleotídeos pirimidínicos ativados (Powner et al. 2009), que evita moléculas de base e de açúcar e sua condensação, produzindo em vez disso as moléculas finais por meio de intermediários que contribuem átomos tanto para a porção de açúcar quanto para a porção de base dos ribonucleotídeos, a relevância da catálise por aminoácidos na produção de D-açúcares ou qualquer outra possível participação de aminoácidos enriquecidos em enantiômeros na síntese de nucleotídeos não é clara. Como a gliceraldeído é uma das moléculas de partida na nova síntese (Fig. 1), a quiralidade no RNA poderia ter surgido por um aporte de gliceraldeído homóquiro, cuja origem poderia ter resultado de processos semelhantes aos discutidos para aminoácidos.
De qualquer forma, também foi demonstrado que, uma vez presente um excesso enantiomérico de aminoácidos, as reações catalisadas por eles poderiam aumentar ainda mais o excesso enantiomérico. Foi demonstrado que, na catálise da síntese de açúcares por aminoácidos, pode ocorrer uma amplificação significativa do excesso enantiomérico (Cordova et al. 2005b). Por exemplo, uma reação mediada pelo aminoácido prolina com um excesso enantiomérico tão baixo quanto 40 % ainda resultou em quase açúcar hexose enantiopuro. A reação catalisada pela prolina com 10 % de excesso enantiomérico forneceu o açúcar com 33 % de excesso enantiomérico.
*****
Em resumo, um cenário plausível para o surgimento da homociralidade na Terra pré-biótica pode ser imaginado com base em todas as descobertas descritas:
1. Origem de excessos enantioméricos locais, ligeiros ou pronunciados, de aminoácidos
2. Caso esses excessos enantioméricos sejam apenas ligeiros, uma amplificação tremenda por equilíbrios entre fase sólida e fase líquida; estes também podem aumentar excessos enantioméricos já elevados
Como vimos no exemplo da síntese de açúcar, a catálise por aminoácidos poderia ter desempenhado um papel na emergência de outros produtos quirais relevantes para a origem da vida, e a amplificação quiral também poderia ter ocorrido durante o processo.
[Se enantiômeros específicos de peptídeos pequenos foram os catalisadores (cf. acima), eles poderiam ter surgido por conjugação de aminoácidos após enriquecimento enantiomérico.]
*****
Uma vez que a homociralidade surge em algumas biomoléculas, ela pode aparentemente impor a homociralidade correspondente às outras. Já vimos isso com a catálise de aminoácidos, mas isso também pode ser válido para a relação entre RNA e aminoácidos. Uma vez que os açúcares D estivessem presentes no RNA, o passo em direção aos aminoácidos L nas proteínas poderia ter sido automático. A síntese proteica requer a aminoacilação do RNA, e é neste passo que os aminoácidos L poderiam ter sido selecionados, como mostra um estudo elegante (Tamura, Schimmel 2004).
No estudo, foi utilizado um minihélice de RNA que recapitula o domínio dentro dos RNAs de transferência que abriga o local de ligação de aminoácidos. Tais minihélices de RNA são consideradas os progenitores dos modernos RNAs de transferência, que apresentam aminoácidos ligados a eles à maquinaria ribossômica que produz proteínas. Os autores demonstraram que o minihélice de RNA era aminoacilado por aminoácidos ativados com uma clara preferência por L- em detrimento de D-aminoácidos. Um sistema de RNA de imagem espelhada mostrou a seletividade oposta.
Três aminoácidos — alanina, leucina e fenilalanina — foram testados, e a seletividade observada foi de 4 vezes. Os autores apontam em uma publicação subsequente, que estuda o mecanismo (Tamura, Schimmel 2006), que tal um efeito de 4 vezes, repetido sob pressão seletiva muitas vezes, pode levar a uma preferência esmagadora por um aminoácido L em um sistema biológico.
Por outro lado, as cadeias peptídicas também poderiam ter adquirido quiralidade independentemente do RNA, veja por exemplo Saghathelian et al. 2001, veja também comunicado de imprensa, Hitz e Luisi 2004, Weissbuch et al. 2004, Plankensteiner et al. 2005. É claro que, qualquer que sejam esses mecanismos em peptídeos, também poderiam ter sido auxiliados por um excesso enantiomérico de aminoácidos, como alcançado nos cenários descritos acima.
Qualquer que tenha sido a sequência exata de eventos na origem da vida, a força cumulativa de todos os dados acima indica que o mistério da quiralidade da origem da vida pode, afinal, ser bem explicado.
*****
A questão da quiralidade, entre outras, tem sido apresentada por criacionistas como um "grande problema" para o conceito de uma origem da vida por causas naturais. Alegadamente, apenas uma intervenção milagrosa de Deus poderia ter resolvido o problema. No entanto, as descobertas acima são um exemplo típico do por que o conceito de Deus dos intervalos não funciona: a ciência fecha rapidamente os intervalos que anteriormente poderiam ter sido considerados reservados para intervenção milagrosa.
Isto é exatamente o que deveria ser esperado se o mundo material for tudo o que existe, ou se o mundo foi criado por um Deus que, como causa primária, escolheu criar através de causas secundárias precisamente aquelas causas naturais que a ciência estuda. De fato, os criacionistas deveriam seriamente perguntar a si mesmos se o seu conceito de Deus não é um conceito que menospreza: o Designer Inteligente como "reparador" que é forçado a quebrar as próprias leis da natureza criadas de vez em quando porque elas são insuficientes para alcançar certas etapas no desenvolvimento do mundo material. De uma perspectiva filosófica teísta, as descobertas reais da ciência sugerem uma ideia muito mais grandiosa de Deus: o Designer que estabeleceu um conjunto elegante e auto-suficiente de leis da natureza que realizam o desenrolar da sua criação induzindo a auto-organização do mundo material. Esta ideia é facilmente compatível com o conceito de Deus de muitas religiões mainstream, incluindo a maioria das cristãs.
Os hiperciclos, como uma organização abrangente de conjuntos autocatalíticos, foram propostos como um modelo para a origem da vida (Eigen e Schuster 1977) e são explicados em Principia Cybernetica Web. No entanto, os hiperciclos complexos só existem como simulações computacionais. Mais de trinta anos após a introdução da hipótese, não há qualquer evidência experimental para hiperciclos complexos, possivelmente pré-bióticos. Isso os torna ainda nada mais do que mera especulação e, embora a hipótese tenha sido um tema recorrente na literatura científica sobre a origem da vida, ela não é mais frequentemente mencionada em publicações recentes no campo.
Até agora, apenas um, muito simples, hiper-ciclo foi gerado há mais de 10 anos no laboratório com dois peptídeos autorreplicantes pelo grupo de Ghadiri (Lee et al. 1997. No entanto, esses autores publicaram uma correção (Lee et al. 1998) onde afirmam: Embora os dados cinéticos sugiram a intermediação de espécies de ordem superior nos processos autocatalíticos, o presente sistema não deve ser referido como um exemplo de hiper-ciclo mínimo na ausência de evidência experimental direta para o acoplamento cruzado auto-catalítico entre replicadores.
Existe apenas um relato sobre um hiper ciclo que ocorre naturalmente (Eigen et al. 1991). Mas este exemplo, relacionado ao ciclo de infecção de um bacteriófago de RNA, obviamente não é pré-biótico, pois o ciclo evoluiu na complexidade dos seres vivos como molde.
Embora pesquisas recentes e empolgantes tenham fornecido cenários plausíveis para a origem da vida e tenham respondido a muitas questões, é claro que muito trabalho de pesquisa ainda precisa ser feito, já que grande parte dos cenários de origem da vida ainda é hipótese. São necessários modelos experimentais que sejam tanto realistas quanto de alguma complexidade apreciável. Se fosse possível, por exemplo, demonstrar que um organismo de RNA primitivo poderia ser construído em laboratório (Szostak et al. 2001), isso seria um passo significativo para frente. Para isso, veja o artigo de Carl Zimmer; ali também é expressa a esperança de que a evolução de tal organismo possa ser observada no laboratório.
(1) Fora de um contexto celular, a evolução do RNA em laboratório é agora rotineira. A engenharia de RNA, de acordo com mecanismos que se assemelham em parte à evolução, demonstrou produzir muitos catalisadores de RNA (veja, por exemplo, Robertson et al. 2004). Certamente, todos os procedimentos sofisticados utilizados em laboratório não estariam disponíveis para um precursor antigo de uma célula. Por outro lado, nada em um tubo de ensaio se assemelha às possibilidades que surgiriam da competição de células replicantes.
(2) A hipótese de que a vida pode ter se originado em fontes hidrotermais parece, em certa medida, atraente também devido ao fato de que sulfetos metálicos, encontrados nessas localidades e capazes de catalisar reações orgânicas simples, ainda são encontrados nos centros catalíticos de enzimas metabólicas centrais, como ferredoxina, succinato desidrogenase e acetil-CoA sintase bacteriana, possivelmente indicando a preservação evolutiva da catálise por sulfetos metálicos primitivos.
(3) Wchtershuser apresenta argumentos teóricos interessantes sobre por que o metabolismo superficial, em oposição a reações em solução, deve abordar a especificidade da reação de maneira suficiente, no entanto, sua perspectiva também foi criticada por Christian De Duve e Stanley Miller em um artigo conjunto (De Duve, Miller 1991). Evidência experimental para a especificidade de uma sequência de reações estritamente confinada a uma superfície mineral (não apenas catalisada por ela, o que permitiria a difusão de produtos intermediários para longe e de volta para superfícies catalíticas) seria difícil de obter, e também as próprias publicações experimentais de Wchtershuser, bem como as de outros que testam seu cenário de metabolismo primeiro, até agora não abordam realmente essa questão central à sua hipótese, mas concentram-se em outros aspectos mais simples de seu modelo.
(4) Vários exemplos de como a oligomerização prebiótica de aminoácidos em pequenos peptídeos poderia ter ocorrido:
a) Via a reação de formação de peptídeos induzida por sal (SIPF), possivelmente em poças, poças de maré ou lagunas (Schwendinger, Rode 1989)
b) Por gás vulcânico (Leman et al. 2004)
c) Em fontes hidrotermais de águas profundas, por ativação com monóxido de carbono (CO) na presença de minerais (Huber et al. 2003)
(5) Altamente específica principalmente em termos de quimiospecificidade; em termos de estereoespecificidade apenas quando aplicável. Esta catálise específica também poderia ocorrer em conjunto com a formação de complexos em superfícies minerais, como as encontradas em fontes hidrotermais de águas profundas, ou moléculas orgânicas poderiam formar complexos com metais solubilizados neste ambiente de alta temperatura e alta pressão (cf. Cody et al. 2000).
Baaske P, Weinert FM, Duhr S, Lemke KH, Russell MJ, Braun D (2007) Acumulação extrema de nucleotídeos em sistemas de poros hidrotermais simulados. Proc. Natl. Acad. Sci. USA 104: 9346-9351 [Pubmed] [Texto Completo Gratuito]
Bada JL, Fegley B Jr, Miller SL, Lazcano A, Cleaves HJ, Hazen RM, Chalmers J (2007) Debating evidence for the origin of life on Earth. Science 315:937-939 [Pubmed]
Bailey J, Chrysostomou A, Hough JH, Gledhill TM, McCall A, Clark S, Menard F, Tamura M (1998) Polarização circular em regiões de formação estelar: implicações para a homociralidade de biomoléculas. Science 281: 672-674 [Pubmed]
Basset M-P (2003) A água de alta pressão é o berço da vida? JPCM 15: 1353-1361 [Resumo, texto completo gratuito disponível lá]
Bokov K, Steinberg SV (2009) Um modelo hierárquico para a evolução do RNA ribossomal 23S. Nature 457: 977-980 [Pubmed]
Breslow R, Levine SM (2006) Amplificação de concentrações enantioméricas sob condições pré-bióticas críveis. Proc. Natl. Acad. Sci. USA 103:12979-12980 [Pubmed] [Texto Completo Gratuito]
Budin I, Bruckner RJ, Szostak JW (2009) Formação de vesículas semelhantes a protocélulas em uma coluna de difusão térmica. J. Am. Chem. Soc. 131: 9628-9629 [Pubmed] [Artigo completo na página de publicações do laboratório de Szostak]
Castro-Puyana M, Salgado A, Hazen RM, Crego AL, Alegre ML (2008) A primeira contribuição da eletroforese capilar para o estudo das origens abióticas da homociralidade: Investigação da adsorção enantioseletiva do ácido 3-carboxi adípico em minerais. Eletroforese 29:1548-1555 [Pubmed]
Chen IA, Roberts RW, Szostak JW (2004) O surgimento da competição entre protocélulas modelo. Science 305:1474-1476 [Pubmed] [Artigo completo na página de publicações do laboratório de Szostak]
Chyba C, Sagan C (1992) Produção endógena, entrega exógena e síntese por impacto de moléculas orgânicas: um inventário para as origens da vida. Nature 355:125-132 [Pubmed]
Chyba CF (2005) Reavaliando a atmosfera primitiva da Terra. Science 308: 962-963 [Pubmed]
Cleaves HJ, Chalmers JH, Lazcano A, Miller SL, Bada JL (2008) Uma reavaliação da síntese orgânica pré-biótica em atmosferas planetárias neutras. Orig. Life Evol. Biosph. 38:105-115 [Pubmed]
Cody GD, Boctor NZ, Filley TR, Hazen RM, Scott JH, Sharma A, Yoder HS Jr. (2000) Compostos de ferro-enxofre carbonilados primordiais e a síntese de piruvato. Science 289:1337-1340 [Pubmed]
Cordova A, Ibrahem I, Casas J, Sunden H, Engquist M, Reyes E (2005a) Neogênese de carboidratos catalisada por aminoácidos: uma transformação antiga plausível. Chemistry 11: 4772-4784 [Pubmed]
Cordova A, Engquist M, Ibrahem I, Casas J, Sunden H. (2005b) Origens plausíveis da homociralidade na neogênese de carboidratos catalisada por aminoácidos. Chem. Commun. (Camb) (2005): 2047-2049 [Pubmed]
Cordova A, Zou W, Dziedzic P, Ibrahem I, Reyes E, Xu Y (2006) Reações aldol intermoleculares assimétricas diretas catalisadas por aminoácidos e pequenos peptídeos. Chemistry 12: 5383-5397 [Pubmed]
Cronin JR, Pizzarello S (1997) Excesso enantiomérico em aminoácidos meteoríticos. Science 275: 951-955 [Pubmed]
De Duve C, Miller SL (1991) Vida bidimensional? Proc. Natl. Acad. Sci. USA 88: 10014-10017 [Pubmed] [Texto Completo Gratuito]
De Duve, C (1995) As origens da vida na Terra. American Scientist 83: 428-437 [Texto completo gratuito]
Eder U, Sauer G, Wiechert R (1971) Novo Tipo de Ciclização Assimétrica para Produzir Esteroide Óptico Ativo CD Estruturas Parciais. Angew. Chem. Int. Ed. Engl. 10:496-497
Eigen M, Schuster P (1977) O hiper-ciclo. Um princípio de auto-organização natural. Parte A: Emergência do hiper-ciclo. Naturwissenschaften 64: 541-565
Eigen M, Biebricher CK, Gebinoga M, Gardiner WC (1991) O hiper-ciclo. Acoplamento da biossíntese de RNA e proteína no ciclo de infecção de um bacteriófago de RNA. Biochemistry 30:11005-11008
Gil R, Silva FJ, Pereto J, Moya A (2004) Determinação do núcleo de um conjunto gênico mínimo bacteriano. Microbiol. Mol. Biol. Rev. 68: 518-537 [Pubmed] [Texto Completo Gratuito]
Glass JI, Assad-Garcia N, Alperovich N, Yooseph S, Lewis MR, Maruf M, Hutchison CA 3rd, Smith HO, Venter JC (2006) Genes essenciais de uma bactéria mínima. Proc. Natl. Acad. Sci. USA 103: 425-430 [Pubmed] [Texto Completo Gratuito]
Glavin DP, Dworkin JP (2009) Enriquecimento do aminoácido L-isovalina por alteração aquosa em corpos parentais de meteoritos CI e CM. Proc. Natl. Acad. Sci. USA 106: 5487-5492 [Pubmed]
Hajos ZG, Parrish DR (1974) Síntese assimétrica de intermediários bicíclicos da química de produtos naturais. J. Org. Chem. 39:1615-1621 [Resumo]
Hanczyc, MM, Fujikawa, SM, Szostak, JW (2003) Modelos experimentais de compartimentos celulares primitivos: Encapsulamento, crescimento e divisão. Science 302:618-622 [Pubmed] [Artigo completo na página de publicações do laboratório de Szostak]
Hanczyc MM, Szostak JW (2004) Vesículas replicantes como modelos de crescimento e divisão de células primitivas. Curr. Opin. Chem. Biol. 8:660-664 [Pubmed] [Artigo completo na página de publicações do laboratório de Szostak]
Hanczyc MM, Mansy SS, Szostak JW (2007) Montagem de Membranas Orientada por Superfície Mineral. Orig. Life Evol. Biosph. 37:67-82 [Pubmed] [Artigo completo na página de publicações do laboratório de Szostak]
Hazen, RM, Filley TR, Goodfriend GA (2001) Adsorção seletiva de aminoácidos L- e D em calcita: implicações para a homociralidade bioquímica. Proc. Natl. Acad. Sci. USA 98:5487-5490 [Pubmed] [Texto Completo Gratuito]
Hazen RM, Boctor N, Brandes JA, Cody GD, Hemley RJ, Sharma A, Yoder Jr HS (2002) Alta pressão e a origem da vida. JPCM 14: 11489-11494 [Resumo]
Hazen RM (2005) Gen-e-sis: A Busca Científica pelas Origens da Vida, Joseph Henry Press
Hitz TH, Luisi PL (2004) Início espontâneo da homociralidade em cadeias de oligopeptídeos geradas na polimerização de aminoácidos de N-carboxianidrido em água. Orig. Life Evol. Biosph. 34:93-110 [Pubmed]
Huang W, Ferris JP (2006) Síntese de um passo e regioespecífica de até 50-mers de oligômeros de RNA por catalise de montmorilonita. J. Am. Chem. Soc. 128: 8914-8919 [Pubmed]
Huber C, Wächtershäuser G (1997) Ácido acético ativado pela fixação de carbono em (Fe,Ni)S sob condições primordiais. Science 276:245-247 [Pubmed]
Huber C, Eisenreich W, Hecht S, Wchtershuser G (2003) Um possível ciclo primordial de peptídeos. Science 301: 938-940 [Pubmed]
Huber C, Wächtershäuser G (2006) ácidos α-hidroxil e α-amino sob possíveis condições de origem da vida hadiana e vulcânica. Science 314:630-632 [Pubmed]
Johnson AP, Cleaves HJ, Dworkin JP, Glavin DP, Laczano A, Bada JL (2008) O experimento de descarga elétrica vulcânica de Miller. Science 322:404 [Pubmed]
Johnston WK, Unrau PJ, Lawrence MS, Glasner ME, Bartel DP (2001) Polimerização de RNA catalisada por RNA: extensão de iniciador precisa e geral modelada por RNA. Science 292: 1319-1325 [Pubmed]
Joyce GF (2002) A antiguidade da evolução baseada em RNA. Nature 418:214-221 [Pubmed]
Kauffman, Stuart. (1993) Origens da Ordem: Auto-organização e Seleção na Evolução, Oxford University Press [Amazon]
Kasting JF, Zahnle KJ, Walker JCG (1983) Fotoquímica do metano na atmosfera primitiva da Terra. Precambrian Res. 20: 121-148 [Resumo]
Klussmann M, Iwamura H, Mathew SP, Wells DH Jr, Pandya U, Armstrong A, Blackmond DG (2006) Controle termodinâmico da amplificação assimétrica na catálise de aminoácidos. Nature 441: 621-623 [Pubmed]
Klussmann M, Toshiko I, White AJP, Armstrong A, Blackmond DG (2007) Emergência da homociralidade em fase de solução via engenharia de cristais de aminoácidos. J. Am. Chem. Soc. 129: 7657-7660 [Pubmed]
Kojo S, Uchino H, Yoshimura M, Tanaka K (2004) A asparagina racêmica D,L causa excesso enantiomérico de outros aminoácidos racêmicos D,L coexistentes durante a recristalização: uma hipótese explicando a origem dos aminoácidos L na biosfera. Chem. Commun. (2004): 2146-2147 [Pubmed]
Koonin EV (2007) Um reator de produção de RNA para a origem da vida. Proc. Natl. Acad. Sci. USA 104: 9105-9106 [Texto Completo Gratuito]
Koonin EV, Novozhilov AS (2009) Origem e evolução do código genético: o enigma universal. IUBMB Life 61:99-111 [Pubmed]
Koonin EV, Martin W (2005) Sobre a origem de genomas e células dentro de compartimentos inorgânicos. Trends Genet. 21:647-654 [Pubmed]
Kuhn WR, Atreya SK (1979) Fotólise de amônia e o efeito estufa na atmosfera primordial da Terra. Icarus 37: 207-213 [Pubmed]
Kun A, Santos M, Szathmary E (2005) Ribozimas reais sugerem um limite de erro relaxado. Nat. Genet. 37:1008-1011 [Pubmed]
Lee DH Severin K, Yokobayashi Y, Ghadiri MR (1997) Emergência da simbiose na autorreprodução de peptídeos através de uma rede hipercíclica. Nature 390:591-594 [Pubmed]
Lee DH Severin K, Yokobayashi Y, Ghadiri MR (1998) Erratum: Emergência da simbiose na auto-replicação de peptídeos através de uma rede hipercíclica Nature 394:101
Leman L, Orgel LE, Ghadiri MR (2004) Carbonyl sulfide-mediated prebiotic formation of peptides. Ciência 306: 283-286 [Pubmed]
McBride JM, Tully JC (2008) Química física: A vida começou a partir de um atrito? Nature 452:161-162 [Pubmed]
Mansy SS, Schrum JP, Krishnamurthy M, Tob S, Treco DA, Szostak JW (2008) Síntese dirigida por molde de um polímero genético em um protocélula modelo. Nature 454:122-125 [Pubmed] [Artigo completo na página de publicações do laboratório de Szostak]
Mansy SS, Szostak JW (2008) Estabilidade térmica de membranas de protocélulas modelo. Proc. Natl. Acad. Sci. USA 105: 13351-13355 [Pubmed] [Texto Completo Gratuito]
Martin W, Russell MJ (2003) Sobre as origens das células: uma hipótese para as transições evolutivas da geoquímica abiótica a procariotos quemoautotróficos, e de procariotos a células nucleadas. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 358:59-83 [Pubmed] [Texto Completo Gratuito]
Miller SL (1953) Produção de aminoácidos sob condições primitivas da Terra possível. Science 117 (1953): 528-529 [PDF]
Morowitz HJ, Kostelnik JD, Yang J, Cody GD (2000) A origem do metabolismo intermediário. Proc. Natl. Acad. Sci. USA 97: 7704-7708 [Pubmed] [Texto Completo Gratuito]
Noorduin WL, Izumi T, Millemaggi A, Leeman M, Meekes H, Van Enckevort WJ, Kellogg RM, Kaptein B, Vlieg E, Blackmond DG (2008) Emergência de um único estado sólido quiral a partir de um derivado de aminoácido quase racêmico. J. Am. Chem. Soc. 130: 1158-1159
Orgel LE (1998) A origem da vida: uma revisão de fatos e especulações. Trends Biochem. Sci. 23: 491-495 [Pubmed]
Orgel LE (2000) Ciclos bioquímicos auto-organizadores. Proc. Natl. Acad. Sci. USA 97: 12503-12507 [Pubmed] [Texto Completo Gratuito]
Orgel LE (2004) Química pré-biótica e a Origem do Mundo do RNA. Crit. Rev. Biochem. Mol. Biol. 39: 99-123 [Pubmed]
Orgel LE (2006) Síntese Geotérmica e Metabolismo. Astrobiology 6: 297-298
Pavlov AA, Kasting JF, Brown LL, Rages KA, Freedman R (2000) Aquecimento estufa por CH4 na atmosfera da Terra primitiva. J. Geophys. Res. 105: 11981-11990 [Pubmed]
Pizzarello S, Weber AL (2004) Aminoácidos pré-bióticos como catalisadores assimétricos. Science 303:1151
Plankensteiner K, Reiner H, Rode BM (2005) Diferenciação estereosseletiva na reação de formação de peptídeos induzida por sal e sua relevância para a origem da vida. Peptides 26:535-541 [Pubmed]
Poole AM (2006) Passar de um mundo de RNA para células modernas ficou um pouco mais fácil. BioEssays 28:105-108 [Pubmed]
Powner MW, Gerland B, Sutherland JD (2009) Síntese de pirimidinas ribonucleotídeos ativadas em condições plausíveis pré-bióticas. Nature 459: 239-242 [Pubmed]
Rajamani S, Vlassov A, Benner S, Coombs A, Olasagasti F, Deamer D (2008) Síntese de polímeros semelhantes ao RNA a partir de mononucleotídeos auxiliada por lipídios. Orig. Life Evol. Biosph. 38:57-74 [Pubmed]
Rees
M (2001) Just Six Numbers: The Deep Forces that Shape the Universe,
Basic Books [Amazon]
Ricardo A, Carrigan MA, Olcott AN, Benner SA (2004)
Minerais de borato estabilizam a ribose. Science 303:196
Robertson MP, Knudsen SM, Ellington AD (2004) Seleção in vitro de ribozimas dependentes de peptídeos para atividade. RNA 10:114127 [Pubmed] [Texto Completo Gratuito]
Robinson R (2005) Jump-Starting a
Cellular World: Investigating the Origin of Life, from Soup to Networks. PLoS
Biol 3(11): e396 [Texto
Completo Gratuito]
Russell MJ, Martin W (2004) The rocky roots of the
acetyl-CoA pathway. Trends Biochem. Sci. 29: 358-363 [PDF]
Sagan C, Chyba C (1997) The early faint sun paradox: organic shielding of
ultraviolet-labile greenhouse gases. Science 276:1217-1221[Pubmed]
Saghatelian A, Yokabayashi Y, Soltani K, Ghadiri MR (2001) A chiroselective
peptide replicator. Nature 409:797-801 [Pubmed]
Schaefer L, Fegley Jr. B (2007) Desgaseificação de material condrito ordinário e algumas de suas implicações para a química de asteroides, planetas e satélites. Icarus 186:462-483 [Resumo] [PDF]
Schwendinger MG, Rode BM (1989) Papel possível do cobre e cloreto de sódio na evolução pré-biótica de peptídeos. Analytical Science 5: 411-444 [Resumo] [Texto Completo Gratuito]
Smith E, Morowitz HJ (2004) Universalidade no metabolismo intermediário. Proc. Natl. Acad. Sci. USA 101: 13168-13173 [Pubmed] [Texto Completo Gratuito]
Smolin L (1999) A Vida do Cosmos, Oxford University Press [Amazon]
Susskind L (2006) A Paisagem Cósmica: Teoria das Cordas e a Ilusão do Design Inteligente, Back Bay Books [Amazon]
Szostak JW, Bartel DP, Luisi PL (2001) Sintetizando a Vida. Nature 409: 387-390 [Artigo completo na página de publicações do laboratório de Szostak]
Szostak JW (2009) Química de sistemas na Terra primitiva. Nature 459: 171-172
Tamura K, Schimmel P (2004) Aminoacilação seletiva por quiralidade de um minihélice de RNA. Science 305:1253 [Pubmed]
Tamura K, Schimmel P (2006) Aminoacilação seletiva por quiralidade de um minihélice de RNA: características mecanísticas e supressão quiral. Proc. Natl. Acad. Sci. USA 103:13750-13752 [Pubmed] [Texto Completo Gratuito]
O Mundo do RNA. (2006) Editado por: Gesteland RF, Czech TR, Atkins JF. Cold Spring Harbor Laboratory Press [Pubmed]
Tian F, Toon OB, Pavlov AA, De Sterck H (2005) Uma atmosfera terrestre primitiva rica em hidrogênio. Science 308: 1014-1017 [Pubmed]
Viedma C (2001) Cristalização enantiomérica de DL-aspártico e DL-glutâmico: implicações para a quiralidade biomolecular na origem da vida. Orig. Life Evol. Biosph. 31: 501-509 [Pubmed]
Viedma C, Ortiz JE, de Torres T, Izumi T, Blackmond DG (2008) Evolução da homociralidade de fase sólida para um aminoácido proteogênico. J. Am. Chem. Soc. 130:15274-15275 [Pubmed]
Wächtershäuser G (1988) Antes das enzimas e dos modelos: teoria do metabolismo de superfície. Microbiol. Rev. 52: 452-484 [Texto Completo Gratuito]
Wächtershäuser G (1990) Evolução dos primeiros ciclos metabólicos. Proc. Natl. Acad. Sci. USA 87: 200-204 [Pubmed] [Texto integral gratuito]
Walker JC, Brimblecombe P (1985) Ferro e enxofre no oceano pré-biológico. Precambrian
Res. 28:205-222 [Pubmed]
Wedyan M, Preston MR (2005) Adsorção seletiva de isômeros de aminoácidos por componentes de sedimentos naturais. Environ. Sci.
Technol. 39:2115-2119 [Pubmed]
Weissbuch I, Bolbach G, Leiserowitz L, Lahav M (2004) Amplificação quiral de oligopeptídeos via polimerização em cristalitos bidimensionais sobre água. Orig. Life Evol. Biosph. 34:79-92 [Pubmed]
Wolf YI, Koonin EV (2007) Sobre a origem do sistema de tradução e do código genético no mundo do RNA por meio da seleção natural, exaptação e subfuncionalização. Biol. Direct 2:14 [Pubmed] [Texto Completo Gratuito]
Zaher HS, Unrau PJ (2007) Seleção de um ribozima RNA polimerase melhorado com
extensão e fidelidade superiores. RNA 13: 1017-1026 [Pubmed] [Texto Completo Gratuito]
Zahnle KJ (1986) Fotoquímica do metano e a formação de ácido cianídrico (HCN) na atmosfera primitiva da Terra. J. Geophys. Res. 91:
2819-2834 [Abstract]
Zhang L, Peritz A, Meggers E (2005) Um ácido nucleico glicólico simples. J. Am.
Chem. Soc. 127:41744175 [Pubmed]
Zhang XV, Martin ST (2006) Impulsionamento das partes do ciclo de Krebs em sentido reverso através da fotoquímica mineral. J. Am. Chem. Soc.
128: 16032-16033 [Pubmed]
Zhu TF, Szostak JW (2009) Crescimento e Divisão Acoplados de Membranas de Protocélulas Modelo. J. Am. Chem. Soc. 131: 5705-5713 [Pubmed] [Artigo completo na página de publicações do laboratório de Szostak]
[Voltar às Perguntas Frequentes sobre Abiogênese]