El Origen de la Vida
Derechos de autor 2010
[Publicado el 31 de octubre de 2006]
Actualización 1 de julio de 2010
Resumen
Incluso las células más simples actualmente vivas contienen cientos de proteínas, la mayoría de las cuales son esenciales para su funcionamiento. Sin embargo, tal complejidad no pudo haber existido en el origen de la vida. Basándose en investigaciones en el campo, aquí se propone cómo, una vez que existió una molécula genética autorreplicante, pudo haber comenzado la vida y cómo se hizo posible la evolución gradual de la complejidad, en contraste con la aparición repentina de la complejidad que los creacionistas afirman que fue necesaria al comienzo de la vida.Se revisan las condiciones para la síntesis de moléculas orgánicas en la Tierra primitiva y se discuten los modelos de genes primero y metabolismo primero. Mientras que el origen de la homociralidad de los aminoácidos y los azúcares ha sido un problema desconcertante durante décadas, los hallazgos recientes proporcionan explicaciones plausibles.
Otros enlaces:
|
Contenido
- Introducción
- Condiciones para la síntesis de moléculas orgánicas en la Tierra primitiva
- ¿Un mundo de ARN primero?
- Acumulación gradual de complejidad
- Minerales y el origen de la vida
- Especificidad de las reacciones químicas y el escenario del metabolismo primero
- Origen de la homociralidad de los aminoácidos y azúcares
- Hiperciclos
- Perspectivas
- Notas
- Referencias
La ciencia nos muestra que el universo evolucionó mediante la autoorganización de la materia hacia estructuras cada vez más complejas. Los átomos, las estrellas y las galaxias se autoensamblaron a partir de las partículas fundamentales producidas por el Big Bang. En las estrellas de primera generación, se formaron elementos más pesados como el carbono, el nitrógeno y el oxígeno. Las estrellas de primera generación envejecidas luego los expulsaron al espacio; nosotros, que estamos constituidos por estos elementos, nacemos literalmente de polvo estelar. Los elementos más pesados nacieron en las explosiones de supernovas. Posteriormente, las fuerzas de la gravedad permitieron la formación de nuevas estrellas y de planetas. Finalmente, en el proceso de evolución biológica desde células diminutas similares a bacterias (el último ancestro común universal, abrev. LUCA) hasta toda la vida en la Tierra, incluidos nosotros los humanos, surgieron formas de vida complejas a partir de las más simples.
Al considerar esta autoorganización de estructuras materiales en el ámbito de la filosofía, uno puede concluir que ocurre ya sea porque las leyes subyacentes de la naturaleza, que deben ser extremadamente especiales para permitirlo (Rees 2001, Smolin 1999, Susskind 2006), simplemente son como son (posiblemente en el contexto de un multiverso) o porque fueron diseñadas por Dios para este propósito. Dado que sabemos que las leyes de la naturaleza son tan autosuficientes que, basándonos en ellas, la complejidad de todo el universo físico evolucionó desde partículas fundamentales, y además, formas de vida complejas desde las más simples durante la evolución biológica, podemos razonablemente extrapolar que también permitirían que la vida misma surgiera espontáneamente, mediante la evolución química de estructuras adecuadas, independientemente de si creemos que estas leyes están diseñadas o no. Por lo tanto, deberíamos esperar un origen de la vida por causas naturales desde tanto la perspectiva filosófica teísta como la atea.
El estudio experimental del origen de la vida arrancó con el experimento de la sopa prebiótica de Miller (Miller 1953, experimento Miller-Urey), que produjo aminoácidos, esenciales para la vida. En las décadas siguientes, se ha realizado una impresionante química sobre los bloques de construcción de la vida, pero durante mucho tiempo muchas preguntas cruciales carecían de respuestas experimentales que pudieran ofrecer esperanza para direcciones futuras firmes. Entre estas se encontraban la síntesis de nucleótidos, la polimerización de nucleótidos a oligonucleótidos, la incorporación de un gen autoreplicante en células simples sobre las cuales la selección natural pudiera actuar, y el origen de la homociralidad de aminoácidos y azúcares. Conceptualmente, el origen del sistema de traducción de proteínas planteó un problema fundamental. Sin embargo, en la última década se ha logrado un progreso significativo en todas esas áreas, aunque los detalles siguen siendo esquemáticos y persisten problemas en muchos temas. Las reacciones propuestas en el modelo del metabolismo primero, que asume el metabolismo, no los genes, en el origen de la vida, también se han vuelto más prometedoras gracias a hallazgos recientes. En general, se puede decir que las piezas del rompecabezas están empezando a encajar de tal manera que la hipótesis científica de un origen espontáneo de la vida a partir de materia no viva ha logrado finalmente plausibilidad a nivel de evidencia experimental.
Aunque la investigación en este campo ahora parece mucho más prometedora que hace solo una década, la ciencia sobre el origen de la vida, en comparación con la ciencia de la evolución biológica, sigue siendo considerablemente menos desarrollada en su poder explicativo. Como señala Richard Robinson (Robinson 2005): «Si le dan a un biólogo una célula, él le dará el mundo. Pero más allá de asumir que la primera célula debió haber surgido de alguna manera, ¿cómo explican los biólogos su emergencia del mundo prebiótico hace cuatro mil millones de años?»
De hecho, es una cosa que sabemos todos los materiales de construcción químicos de la vida, y que el funcionamiento de la vida puede explicarse completamente por su colaboración en un sistema extremadamente complejo. Sin embargo, es algo totalmente distinto cómo, en el origen de la vida, pudieron formar por sí mismas una organización inicial paso a paso (a través de cualquier proceso intermediario y bloque de construcción). A primera vista, la evolución desde LUCA, un precursor de las bacterias, hasta los humanos puede parecer un juego de niños en comparación: comenzó desde una maquinaria bioquímica ya tremendamente compleja y completamente autosuficiente y, poco a poco, simplemente la hizo aún más compleja.
No se puede perder de vista que las células más elementales que conocemos actualmente, que no dependen permanentemente del metabolismo del huésped, la bacteria Mycoplasma genitalium, tienen 482 genes codificantes de proteínas (la mayoría de las bacterias, como E. coli, codifican para más de 2000 proteínas diferentes), de las cuales, según el estudio experimental más exhaustivo hasta la fecha (Glass et al. 2006), las esenciales son 387. El estudio hipotético probablemente más preciso (Gil et al. 2004) sitúa el número mínimo de genes en 206. Todas las proteínas producidas a partir de estos genes están involucradas en un laberinto de vías de metabolismo, replicación, así como en la construcción y mantenimiento de la estructura, lo cual es de una complejidad atónita.
De hecho, ¿de qué otra manera que a través de una cantidad mínima de complejidad podría incluso una célula primitiva haber satisfecho las demandas básicas mencionadas anteriormente? ¿Cómo podría haber surgido por sí sola una red tan vastamente compleja de más de 200 proteínas? Uno podría preguntar: ¿no tendría que haber surgido de una sola vez? Sin embargo, la evidencia sugiere que toda esta complejidad pudo haber evolucionado, paso a paso, a partir de comienzos muy simples.
2. Condiciones para la síntesis de moléculas orgánicas en la Tierra primitiva
Esencial para el origen espontáneo de la vida fue la disponibilidad de moléculas orgánicas como bloques de construcción. El famoso experimento del caldo prebiótico de Stanley Miller (Miller 1953, experimento Miller-Urey) había demostrado que los aminoácidos, los bloques de construcción de las proteínas, surgieron junto con otras pequeñas moléculas orgánicas de forma espontánea al reaccionar una mezcla de metano, hidrógeno, amoníaco y agua en un aparato de descarga eléctrica. Se asumía que estas condiciones simulaban las de la Tierra primitiva. Ya en 1922, Oparin había propuesto que la Tierra primitiva tenía una atmósfera reductora (en su obra clásica El Origen de la Vida de 1936, amplió estas ideas). Las observaciones de Júpiter y Saturno habían mostrado que contenían amoníaco y metano, e inferían la presencia de grandes cantidades de hidrógeno allí también (ahora se sabe que el hidrógeno es el componente atmosférico principal de estos planetas). Estas atmósferas reductoras de los planetas gigantes se consideraban restos capturados de la nebulosa solar y se asumía por analogía que la atmósfera de la Tierra primitiva había sido similar.
Se sugirió que solo en una atmósfera reductora como esta, la síntesis de moléculas orgánicas, también azúcares y bases orgánicas, bloques de construcción de nucleótidos, habría sido posible en grandes cantidades (Chyba y Sagan 1992).
Investigaciones posteriores pusieron en duda la existencia de una atmósfera reductora y sugirieron una atmósfera neutra en su lugar; véase también (Chyba 2005), el artículo complementario a Tian et al. (véase más abajo).
Sin embargo, nuevos cálculos indican que el hidrógeno escapó de la atmósfera primitiva a un ritmo mucho más lento de lo que se pensaba anteriormente, dando lugar a una atmósfera donde el hidrógeno era un componente principal (aproximadamente 30 %) y que, por lo tanto, era altamente reductora (Tian et al. 2005, véase también comunicado de prensa). Los autores midieron la producción de moléculas orgánicas mediante fotólisis UV bajo esas condiciones, y concluyen que a 1010kg/año habría sido órdenes de magnitud mayor que el ritmo de la síntesis de compuestos orgánicos en sistemas hidrotermales o la entrega exógena de compuestos orgánicos a la Tierra primitiva.
Otro estudio reciente también apoya una atmósfera reductora temprana. Condritas son material primitivo de la nebulosa solar y se cree generalmente que son los bloques de construcción de la Tierra y otros planetas rocosos, asteroides y satélites. Durante y después de la formación planetaria, los gases escapan del material condritico debido a altas temperaturas y presiones. Cálculos sistemáticos y detallados sobre qué gases debieron ser muestran que son principalmente hidrógeno, metano y amoníaco altamente reductores —los mismos gases que en los experimentos tipo Miller-Urey (Schaefer y Fegley 2007, véase también comunicado de prensa). La composición de los gases varió con la temperatura solo en un grado moderado, y se encontró que era en gran medida independiente de la presión real bajo la cual pudo haber ocurrido la desgasificación, lo que parece apoyar la robustez de las conclusiones.
Los autores mencionan que se había encontrado que una atmósfera reductora de metano y amoníaco es extremadamente vulnerable a la destrucción por la luz UV del sol (Kuhn y Atreya 1979, Kasting et al. 1983). Sin embargo, también señalan que los desarrollos recientes sugieren que una atmósfera reductora es más estable de lo que se creía anteriormente:
1) Se encontró que la fuga de hidrógeno desde la atmósfera de la Tierra era menos eficiente de lo que se pensaba anteriormente (refiriéndose al estudio de Tian et al. mencionado anteriormente).
2) Las observaciones de la atmósfera de Titán, la luna de Saturno, que está compuesta principalmente de metano y nitrógeno, muestran que los aerosoles de hidrocarburos producidos fotoquímicamente forman una capa de niebla en la atmósfera superior que protege la atmósfera inferior de la destrucción fotoquímica. Una capa de niebla como esta también podría haberse producido en la Tierra primitiva a partir del metano y el amoníaco expulsados (Zahnle 1986, Sagan y Chyba 1997, Pavlov et al. 2000).
Sin embargo, incluso si la atmósfera de la Tierra primitiva habría sido neutra en lugar de reductora, nuevos datos sugieren que la síntesis eficiente de aminoácidos habría sido posible también bajo estas circunstancias. El grupo de Jeffrey Bada demostró que, en contra de informes anteriores, cantidades significativas de aminoácidos se producen a partir de mezclas de gases neutros bajo condiciones adecuadas (Cleaves et al. 2008). La tamponización con carbonato de calcio (un material rocoso común) previno la disminución del pH causada por los nitrito y nitrato producidos en el aparato de descarga eléctrica utilizado, aumentando drásticamente el rendimiento. Además, la adición de inhibidores de oxidación previno la degradación oxidativa de aminoácidos por nitrito y nitrato, aumentando aún más el rendimiento. Inhibidores de oxidación como los iones ferroso podrían haber estado en exceso sobre el nitrito/nitrato en la Tierra primitiva (Walker y Brimblecombe 1985). El grupo de Bada también reanalizó muestras de los experimentos de descarga eléctrica de Miller de los años 1950 que simulaban erupciones volcánicas ricas en vapor de agua. Tales erupciones habrían liberado gases reductores. En estas muestras, los aminoácidos eran más variados que en el experimento clásico de Miller, y los rendimientos eran comparables o incluso superiores, indicando que incluso si la atmósfera de la Tierra hubiera sido neutra, la síntesis prebiótica localizada podría haber sido efectiva (Johnson et al. 2008, véase también artículo de noticias).
Por supuesto, si la vida surgió en fuentes hidrotermales de aguas profundas (véase más abajo), la composición de la atmósfera primitiva de la Tierra se volvería en gran parte irrelevante. En cierto modo, esto también es cierto para los bloques de construcción orgánicos entregados a la Tierra por partículas de polvo interestelar y en meteoritos carbonáceos.
En una atmósfera reductora, cuya probabilidad se ve favorecida por los hallazgos discutidos anteriormente, la concentración de compuestos orgánicos en el océano prebiótico pudo haber sido relativamente alta (De Duve y Miller 1991, y las referencias allí contenidas). Además, localmente, una sopa prebiótica pudo haberse concentrado enormemente mediante procesos tan simples como, por ejemplo, la evaporación en charcos o lagos someros, posiblemente con ciclos a largo plazo de humedad/seca. Debe tenerse en cuenta al evaluar todos los escenarios químicos que, debido a su naturaleza, el origen de la vida debió ser un evento muy local; esto también es importante para el tema del origen de la homociralidad de los aminoácidos y los azúcares, véase a continuación.
Ahora está ampliamente aceptado que en el origen de la vida no existía el actual sistema de ADN/(ARN)/proteína para la información genética por un lado y la catálisis, regulación y función estructural por el otro. Sería una pregunta sin sentido, ¿qué vino primero, la proteína o el ADN? La catálisis proteica sin información genética, que le permite mantenerse y propagarse, no es suficiente a largo plazo, y la información genética del ADN sin catálisis, necesaria para la función de la vida, también sería inútil.
En cambio, se asume que el ARN actuó como precursor tanto de proteínas como de ADN, en el sentido de que puede servir tanto como catalizador (como las enzimas proteicas) como portador de información genética. Incluso en la célula moderna, los ribozimas (ARNs catalíticos) aún desempeñan un papel vital, aunque limitado. En el ribosoma, la síntesis de las cadenas peptídicas de las proteínas a partir del código de ARN se lleva a cabo mediante ribozimas. También catalizan el empalme del ARN.
La hipótesis de que un llamado Mundo de ARN estuvo involucrado en las etapas evolutivas tempranas de la vida es ahora una visión casi universalmente aceptada (Joyce 2002, Orgel 2004, The RNA World 2006). ¿Podría este Mundo de ARN haber estado en el origen último de la vida? Esta es actualmente aún una pregunta abierta. El sistema de ARN puede ser demasiado complejo para haber surgido sin síntesis por un precursor genético o un metabolismo previo sin enzimas (opciones discutidas a continuación). Sin embargo, aunque todavía existen problemas sustanciales, ahora hay buenas pistas para procesos simples y espontáneos en la Tierra primitiva tanto para la síntesis de nucleótidos como para su concatenación en oligonucleótidos.
Durante mucho tiempo, la síntesis de monómeros de ARN bajo condiciones prebióticas parecía ser un problema fundamental, ya que la condensación de azúcar (ribosa) y nucleobase (purinas y pirimidinas) no funciona (Orgel, 2004). La síntesis prebiótica de purinos ribonucleótidos sigue siendo poco clara, sin embargo, recientemente se ha logrado un avance en lo que respecta a la síntesis de monómeros de pirimidina ribonucleótidos (que incorporan citosina y uracilo). Ahora parece estar resuelto en principio, de una manera completamente inesperada. El estudio del grupo de John Sutherland (Powner et al. 2009) muestra cómo la naturaleza pudo haber ensamblado espontáneamente monómeros de pirimidina ribonucleótidos a partir de moléculas plausibles prebióticamente a través de intermediarios que aportan átomos tanto a la porción de azúcar como a la de base de los ribonucleótidos, evitando así por completo un paso de condensación de azúcar y base (Fig.1). Véase también Nature News para el impacto de estos hallazgos. Aunque aún queda por encontrar una buena vía para la síntesis de purinos ribonucleótidos (que incorporan adenina y guanina), Jack Szostak argumenta en un comentario que acompaña al artículo (Szostak 2009) que es precisamente porque este trabajo abre tantas nuevas direcciones para la investigación que se mantendrá durante años como uno de los grandes avances en química prebiótica.
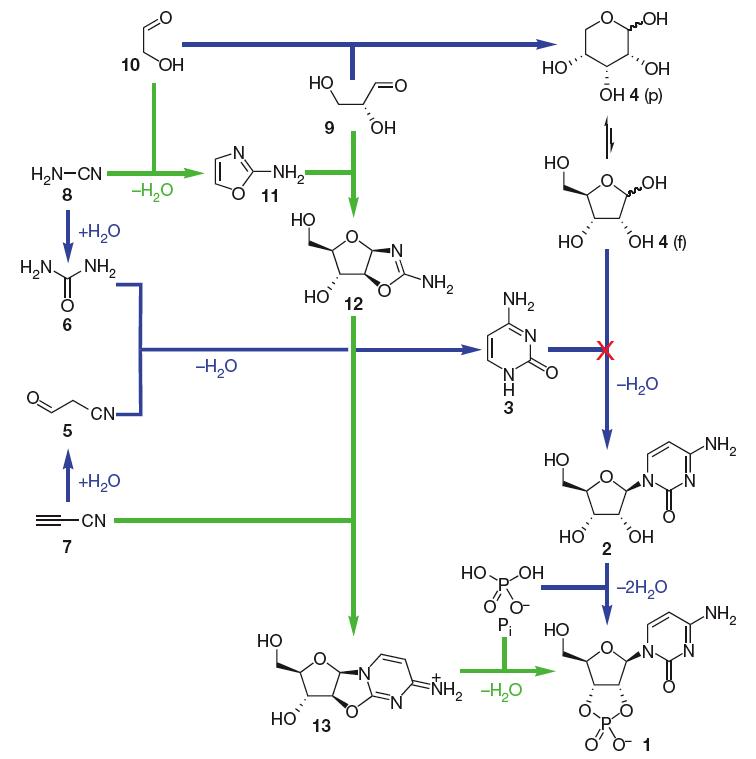
Figura 1. Ensamblaje de pirimidinas ribonucleótidas. Síntesis previamente asumida de b-ribocitidina-2,3-cíclico fosfato 1 (azul; nota el fallo del paso en el que se propone que la citosina 3 y la ribosa 4 se condensan juntas) y la nueva síntesis exitosa descrita aquí (verde). p, piranosido; f, furanosido. De (Powner et al. 2009). Reproducido con permiso de Macmillan Publishers Ltd.
(5 = cianacetaldehído, 6 = urea 6, cianacetileno 7, 8 = cianamida, 9 = gliceraldehído, 10 = glicolaldehído, 11 = 2-amino-oxazol, 12 = pentosa amino-oxazolina, derivado de arabinosa, 13 = anhidroarabinonucleósido)
En cuanto a la concatenación de nucleótidos para formar oligonucleótidos, también hay avances. La polimerización de monómeros de ARN químicamente activados puede tener lugar en las superficies minerales de la arcilla montmorillonita, generando cadenas poliméricas de hasta 50-meros (Huang y Ferris 2006). Los monómeros de ribonucleótidos pirimidínicos de la nueva síntesis (Powner et al. 2009) también están activados (contienen fosfato cíclico), lo que podría permitir una polimerización similar.
El grupo de David Deamer ha demostrado que la síntesis de polímeros similares al ARN puede incluso ocurrir a partir de mononucleótidos no activados dentro de vesículas de fosfolípidos, debido al potencial químico de condiciones anhidras e hidratadas fluctuantes, con el calor proporcionando energía de activación durante la rehidratación (Rajamani et al. 2008). Tales condiciones podrían haber existido alrededor de manantiales termales en la Tierra prebiótica. Los lípidos también proporcionan un microambiente estructuralmente organizador que impone orden a los mononucleótidos. En este montaje experimental, se pueden formar oligómeros de hasta 100 nucleótidos de forma no enzimática. Todavía queda por ver si las vesículas de ácidos grasos plausibles prebióticamente podrían tener el mismo efecto sobre la síntesis de ARN (con esto, una molécula de ARN autorreplicante también habría estado preempacada para una evolución ulterior, cf. abajo). La polimerización efectiva de monómeros que están activados podría ser ayudada por un microambiente estructuralmente organizador dentro de las vesículas también.
Sin embargo, aunque estas reacciones producen polímeros similares al ARN, aún no resuelven el problema de la concatenación estereoespecífica 3-5 de los monómeros (Orgel 2004), presente en todos los organismos vivos. Tanto la síntesis asistida por lípidos como la polimerización sobre montmorillonita producen mezclas de enlaces 2-5 y 3-5. Sin embargo, en este último caso existe una preferencia hacia los enlaces 3-5 (hasta el 74 %) lo cual es prometedor. Los autores del estudio (Huang y Ferris 2006) argumentan que una mayor proporción de enlaces 3-5 podría haber resultado en una evolución más rápida hacia todos los enlaces fosfodiéster 3-5 durante la replicación de oligómeros, sugiriendo que quizás no fue necesaria la especificidad completa de los enlaces 3-5 dentro de la primera molécula de ARN replicante. Este escenario no sería imposible, dado que el ARN suele formar estructuras secundarias más irregulares que el ADN con su doble hélice, para el cual la estereoespecificidad absoluta de la síntesis es obligatoria. En el estudio, diferentes tipos de activación química de los monómeros de ARN dieron lugar a diferentes preferencias para la concatenación 3-5. Queda por ver cuán específica será la concatenación de monómeros en enlaces 3-5 a partir de la activación con fosfato cíclico, el tipo de activación reportado en la nueva síntesis de monómeros de ribonucleótidos pirimidínicos (Powner et al. 2009).
Debido a las dificultades de la síntesis de nucleótidos de ARN y oligonucleótidos, que sin embargo parecen ahora considerablemente menos severas que hace solo unos años, dado los nuevos hallazgos, se han propuesto varias alternativas al ARN como el primer sistema genético que podrían haber precedido a él. El ácido nucleico peptídico (PNA), el ácido nucleico treosa (TNA) y los péptidos en los que los aminoácidos D- y L alternan que incorporan bases estándar de ácido nucleico (ANAs) se discuten en (Orgel 2004). Un sistema aún más simple es el ácido nucleico glicerol (GNA, véase Zhang et al. 2005). Una idea altamente interesante y químicamente atractiva, pero aún no probada, es el Mundo PAH (PAH = hidrocarburos aromáticos policíclicos), desarrollado por S. Platts y descrito en (Hazen 2005) y también en Wikipedia y en pahworld.com.
Otras dificultades son que aún no se ha encontrado un ribozima (ARN catalítico) que pueda copiarse por completo hasta ahora, un ribozima de 200 bases puede copiar alrededor de 20 bases de su secuencia con alta fidelidad (Zaher y Unrau 2007), una mejora respecto a (Johnston et al. 2001). Se ha propuesto una solución para el problema de copiar una secuencia larga de ribozima en Szostak et al. (2001); los experimentos tendrán que demostrar si es factible. Además, el formidable problema de separar el producto de doble cadena de la reacción de copia para permitir una segunda ronda de copiado seguiría por resolverse (Orgel 2004). Los hallazgos recientes sugieren que este problema podría abordarse mediante ciclos térmicos, análogo a la reacción en cadena de la polimerasa (PCR), lo cual podría haber sido una posibilidad bajo condiciones prebióticas plausibles (Mansy y Szostak 2008, Baaske et al. 2007).
4. Acumulación gradual de complejidad
Asumamos el plausible escenario de que el ARN fue sintetizado directamente, véase arriba, de modo que a partir de un gran pool de ARNs aleatorios pudiera surgir una molécula de ARN autorreplicante, o que dicha síntesis fue realizada por un sistema genético/catalítico precursor (posiblemente en la superficie de minerales, cf. Orgel 2004). Dado que los ácidos grasos podrían haber estado disponibles en el ambiente (Hanczyc et al. 2003, Orgel 2004), una membrana primitiva de ácido graso podría haber rodeado a las primeras moléculas de ARN autorreplicantes (debido a sus propiedades moleculares, los ácidos grasos pueden formar vesículas espontáneamente); esto no habría permitido el paso de los polímeros de ARN para que permanecieran juntos, pero habría permitido el paso de los nucleótidos mucho más pequeños, suministrados desde la síntesis prebiótica espontánea o desde un sistema genético/catalítico precursor. Tal membrana habría tenido características diferentes de semipermeabilidad que las membranas lipídicas modernas, donde gran parte del transporte de moléculas está regulado a través de canales proteicos.
El grupo de Jack Szostak ha realizado estudios extensos y plausibles que demuestran que estas vesículas de ácidos grasos, como contenedores para el ARN, habrían permitido el crecimiento y la replicación simplemente mediante mecanismos fisicoquímicos, hasta que una maquinaria de membrana más sofisticada, dirigida por la propia célula y más parecida a lo que se encuentra en los organismos actuales, habría tomado su lugar (Hanczyc et al. 2003, Chen et al. 2004, Hanczyc y Szostak 2004, Zhu y Szostak 2009).
Mientras que en estudios anteriores (Hanczyc et al. 2003, Hanczyc y Szostak 2004) se requerían condiciones más extremas y fuerzas brutas para la división de vesículas, lo que también conducía a la pérdida de una fracción sustancial del contenido de las vesículas, un nuevo estudio (Zhu y Szostak 2009) muestra una solución a estos problemas. Utiliza vesículas multilamelares (vesículas con varias capas de membrana lipídica) que se forman espontáneamente por la rehidratación de películas de ácidos grasos o por la acidificación de una solución concentrada de micelas de ácidos grasos. Una vez formadas las vesículas multilamelares, la incorporación espontánea adicional de micelas de ácidos grasos en ellas provoca, mediante un mecanismo inesperado, la formación de vesículas fuertemente alargadas y en forma de hilo. Tras someterlas a fuerzas de cizallamiento suaves, estas se dividen en varias hijas pequeñas, nuevamente redondas, que preservan bien el contenido de ARN.
El grupo de Szostak también ha demostrado que los nucleótidos pueden atravesar vesículas basadas en ácidos grasos plausibles prebióticamente y que la copia de plantilla no enzimática de una plantilla de oligo dC DNA de modelo puede tener lugar dentro de ellas (Mansy et al. 2008), lo cual, en conexión con los estudios sobre el crecimiento y división de vesículas, revela en principio cómo pudo haber funcionado un protocelula heterótrofo (Fig. 2). Además, mostraron (Mansy y Szostak 2008) que las membranas de modelo plausibles prebióticamente son sorprendentemente termoestables, permitiéndoles tolerar al menos períodos cortos de temperaturas de hasta 100C. El ciclo térmico podría haber sido posible cerca o dentro de la superficie de chimeneas hidrotermales o manantiales calientes (para la convección térmica dentro de poros hidrotermales, véase también el estudio mencionado a continuación por Baaske et al. 2007). Esto podría resolver el espinoso problema de separar el producto de doble cadena de la reacción de copia para una replicación adicional. El ciclo térmico podría haber permitido esta separación de productos de copia (y mayor absorción de nucleótidos, véase Mansy y Szostak 2008) a altas temperaturas, y la copia a temperaturas más bajas, análogo a la reacción en cadena de la polimerasa (PCR).
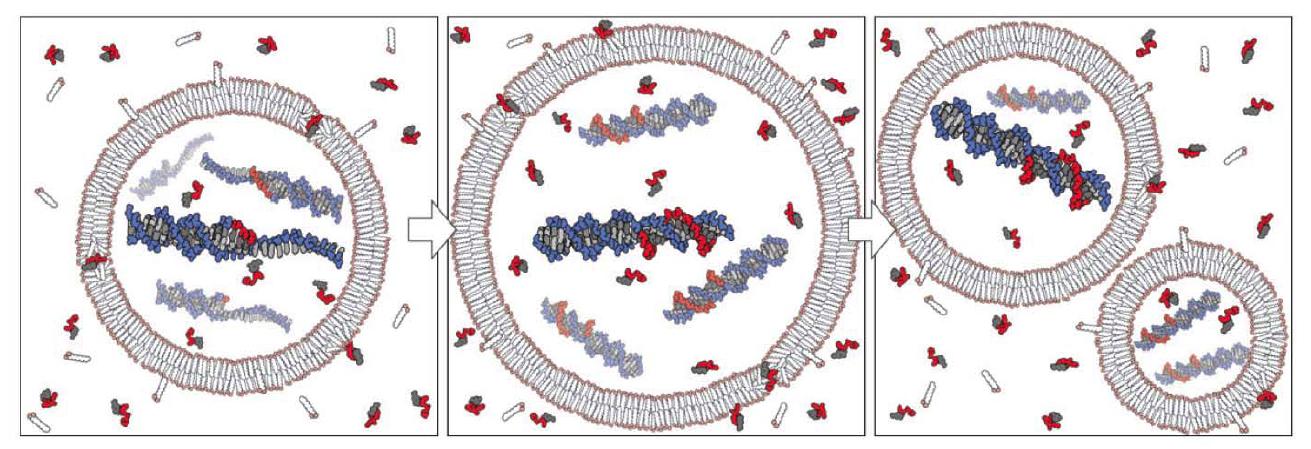
Figura 2. Modelo conceptual de una protocélula heterotrófica. El crecimiento de la membrana de la protocélula resulta de la incorporación de anfifilos suministrados por el medio ambiente, mientras que la división puede ser impulsada por fuerzas físicas intrínsecas o extrínsecas. Los nucleótidos activados suministrados externamente atraviesan la membrana de la protocélula y actúan como sustratos para la copia de las plantillas internas. La replicación completa de la plantilla seguida de la segregación aleatoria del material genético replicado conduce a la formación de protocélulas hijas. De (Mansy et al. 2008). Reproducido con permiso de Macmillan Publishers Ltd.
Extrapolando de todos los datos anteriores, dentro de las vesículas de ácidos grasos, la primera molécula de ARN autorreplicante podría haber comenzado a copiarse a sí misma. Durante la copia, varias cosas habrían sido posibles. Las copias de alta fidelidad habrían producido la misma molécula autorreplicante. Las copias con errores habrían resultado mayoritariamente en ARN no funcional, pero en una minoría de casos, habrían producido ARN que se copiaba más rápido. Se ha demostrado (Chen et al. 2004, véase también artículo de noticias) que los sistemas de ARN/vesículas que contienen más material genético (lo que habría resultado de una replicación de ARN más rápida) desarrollan más tensión interna que las vesículas vecinas que no contienen tanto ARN, y extraen material de membrana de ellas. Importantly, esto habría permitido la selección natural de vesículas por competencia incluso en ausencia de la capacidad de sintetizar sus propios componentes de membrana y, por lo tanto, de controlar directamente su propio crecimiento. Así, por primera vez, un sistema habría tenido la capacidad de experimentar evolución darwiniana mediante la selección natural actuando sobre la variación. Esto habría sido una propiedad emergente nueva y crucial que surgió en la transición de la no vida a la vida.
Una pequeña porción de otros errores de copia (de nuevo, la mayoría de los errores más sustanciales probablemente habrían resultado en moléculas no funcionales, pero esas habrían sido filtradas por la selección natural) podría haber llevado a moléculas de ARN con otras propiedades catalíticas aún diferentes a la función de copia. Una nueva propiedad podría haber permitido al sistema ARN/vesícula competir aún mejor por los recursos: exactamente como en el caso de las moléculas de ARN que presentan la mejor función de copia, el ARN habría evolucionado (1). La función de copia de las moléculas progenitoras probablemente habría actuado sobre estas moléculas hijas también (como una enzima de ARN polimerasa que copia cualquier ARN). Los sistemas ARN/vesícula que tenían las moléculas de ARN alteradas con la nueva función beneficiosa, además de retener el ARN con la función de copia, habrían sido favorecidos por la selección natural. Finalmente, a través de la reiteración de tales procesos, una serie de nuevas propiedades catalíticas podría, por ejemplo, haber permitido que el pool de ARN dentro de las vesículas comenzara a producir sus propios nucleótidos. ¿Sería entonces autosuficiente? En cierto grado, sí. ¿Podría esto haber sido la primera célula primitiva? ¿Por qué no?
Es simplemente que, en este escenario, el metabolismo inicial habría sido mucho más simple que el de hoy: entre otras cosas, el metabolismo energético podría haber sido reemplazado por el paso de bloques de construcción activados para moléculas desde el entorno exterior hacia la vesícula (en un sentido, proporcionando un sustituto preliminar para la producción moderna de ATP, una posibilidad dada el metabolismo simple), y el metabolismo lipídico, la construcción de la estructura de la membrana y su regulación durante la replicación, habría sido reemplazado por vesículas simples que obedecen claramente las fuerzas fisicoquímicas.
En otras palabras: la célula habría dependido más del mundo exterior, pero por lo que estaba haciendo, era en cierto grado autosuficiente (los organismos actuales también necesitan, por supuesto, sustento del mundo exterior, en forma de nutrientes). Por otra parte, la dependencia de la célula del mundo exterior también habría sido posible de una manera más inmediata. Una célula moderna no puede, por ejemplo, utilizar ácidos grasos del exterior de una manera en que se incorporen directamente como elementos de membrana.
Anteriormente preguntamos: ¿Cómo podría haber surgido por sí sola una red compleja de más de 200 proteínas esenciales, como la que se encuentra en las células más elementales de hoy en día?
La clave para responder a esta pregunta parece residir en la combinación de las dos atributos anteriores en la célula primitiva: una dependencia más directa del mundo exterior que la célula moderna, pero también una mayor capacidad para mostrar dicha dependencia al aceptar bloques de construcción moleculares como tales sin tener que convertir nutrientes en ellos. Partiendo de estas características y avanzando gradualmente desde allí, la evolución efectivamente pudo haber facilitado que el sistema celular se dirigiera hacia una mayor complejidad.
Quizás la síntesis de lípidos, en una forma precursora de la síntesis moderna, pudo haber hecho que el sistema fuera más independiente. El sistema de ARN pudo haber, poco a poco, inventado la síntesis de proteínas, como se mencionó; los ribosomas modernos aún contienen ribozimas (ARN catalítico) que catalizan la formación de enlaces peptídicos que finalmente dan lugar a proteínas. En un estudio convincente (Wolf y Koonin 2007), los autores proponen un modelo paso a paso para el origen del sistema de traducción de proteínas, en el que cada paso confiere una ventaja distinta a un conjunto de elementos genéticos coevolutivos. El objetivo del desarrollo de la traducción no habría sido necesario, una visión de futuro que la evolución no tiene. La causa inicial para la emergencia de la traducción habría sido la capacidad de los aminoácidos y los péptidos para estimular reacciones catalizadas por ribozimas (para péptidos demostrados experimentalmente, véase Robertson et al. 2004). Aunque resulte que varios pasos en la evolución de la traducción probablemente hayan sido diferentes del modelo propuesto, el estudio demuestra claramente que no hay nada en la emergencia del sistema de traducción que represente un caso de complejidad irreducible, incapaz de ser objeto de una evolución darwiniana paso a paso.
Un estudio reciente (Bokov y Steinberg 2009), basado en el análisis de la interdependencia de los elementos de la macromolécula, muestra una evolución paso a paso del ribosoma a nivel estructural, comenzando desde un núcleo muy pequeño, el centro de transpeptidación peptídica. Se añadieron capas adicionales desde el exterior una por una, hasta emerger la actual macromolécula compleja. Cada una de estas capas está conectada estructuralmente solo con la respectiva anterior y podría eliminarse en el modelo sin destruir la integridad de las capas internas (como pelar una cebolla), demostrando que no era necesario que toda la molécula emergiera de una vez. Esto establece la falta de complejidad irreducible del mecanismo de traducción desde otra perspectiva.
Finalmente, el metabolismo complejo pudo haberse logrado y la transición al mundo moderno de ADN/(ARN)/proteína. El dualismo ADN/proteína por supuesto es una fuente de complejidad en sí mismo, algo que falta en un organismo con solo ARN.
¿Qué hay del difícil problema de un genoma que contiene todos los genes juntos? Podría haber sido que en las primeras células primitivas los ARNs se unieron por accidente paso a paso, uno a uno, para formar un precursor de genoma y que cada uno de estos pasos conferiera una ventaja en la selección natural frente a células competidoras, ya que los genes no se habrían perdido ya durante la división celular, y la replicación se habría sincronizado. Con el tiempo, un genoma completo de ARN pequeño potencialmente podría haber organizado a sí mismo de esta manera, hasta que los mecanismos para la expansión interna, como los que se encuentran en los genomas modernos, pudieran haberse encargado, por ejemplo, la duplicación génica y la variación del gen duplicado.
Ciertamente, la lectura de una cadena de ARN unido como varios genes individuales habría requerido que el mecanismo celular primitivo hubiera encontrado una manera de reconocer el inicio y el final de una secuencia. Las regiones promotoras y los codones de inicio/fin, en la forma en que se utilizan en los genes codificadores de proteínas, no habrían estado presentes en un organismo de ARN primitivo.
Si bien la falta de fidelidad de replicación habría sido un problema en los genomas de ARN primordiales, el análisis de estudios experimentales de mutación sobre ribozimas indica que un genoma de ARN podría haber crecido hasta alcanzar un tamaño de 100 genes (Kun et al. 2005, Poole 2006). El genoma de ARN podría haber sido reemplazado, poco a poco, por un genoma de ADN, una ventaja seleccionable que las células primordiales habrían encontrado por azar.
En cuanto al código genético universal: hay evidencia de que, hasta el último ancestro común, el código genético fue seleccionado en gran medida para minimizar errores, por lo que no es arbitrario. Sin embargo, en parte probablemente también sea un accidente congelado, ya que la selección por esta característica parece no ser máxima (Koonin y Novozhilov 2009).
En resumen, basándonos en los datos disponibles, el origen espontáneo de la vida como células simples que contienen un polímero genético único, sobre el cual podría actuar la selección natural, es factible. Una transición evolutiva gradual desde estas hacia una complejidad celular común habría sido posible.
5. Minerales y el origen de la vida
Los minerales probablemente proporcionan la clave para resolver una serie de cuestiones relacionadas con el origen de la vida. Como se ha discutido, se ha demostrado que catalizan la polimerización de moléculas similares a nucleótidos (Huang y Ferris 2006, Orgel 2004). También se ha demostrado que permiten la síntesis prebiótica en pureza suficiente de un componente de nucleótido que había resultado elusivo: los minerales de borato estabilizan la ribosa (Ricardo et al. 2004, véase también artículo de noticias; sin embargo, para una síntesis de monómeros de ribonucleótidos sin intermediario de azúcar, véase arriba). La formación de vesículas de ácidos grasos también se ve ayudada por los minerales, y las partículas minerales podrían haber terminado dentro de las vesículas y allí exhibir propiedades catalíticas (Hanczyc et al. 2003, Hanczyc et al. 2007).
En un escenario hipotético diferente, los minerales también desempeñan un papel interesante. En lugar de en una sopa prebiótica acuosa en o cerca de la superficie de la Tierra, se ha hipotetizado que la vida pudo haber comenzado en las profundidades del océano, en el entorno único de las fuentes hidrotermales de aguas profundas. Se ha demostrado que las moléculas orgánicas relevantes pueden sintetizarse en reacciones que involucran gases y minerales presentes en esos sitios, con CO y otras pequeñas moléculas que contienen carbono como fuente de carbono. Estas reacciones requieren las altas temperaturas, o una combinación de altas temperaturas y presiones, encontradas en las fuentes hidrotermales de aguas profundas. Para citar algunos ejemplos: la reacción de CH3SH con CO en presencia de FeS/NiS da como resultado ácido acético activado, o una mezcla de H2S, FeS/NiS y CO (como única fuente de carbono) produce ácido acético (Huber, Wchtershuser 1997). La piruvato se forma a partir de CO en presencia de noniltiol y FeS (Cody et al. 2000). Los ácidos alfa-hidroxílicos y alfa-amino se sintetizan a partir de CO, ligandos cianuro y ligandos metiltio tras la catálisis por precipitados de Fe-Ni, en presencia de hidróxido de calcio o magnesio (Huber, Wchtershuser 2006). Para la crítica de las condiciones en el último estudio, sin embargo, y la respuesta, véase (Bada et al. 2007).
Además, en agua a alta presión y alta temperatura, como la que se encuentra en las fuentes hidrotermales de aguas profundas, las moléculas orgánicas, una vez formadas, muestran un nivel de reactividad química (aunque no siempre particularmente específica) que generalmente solo se observa en ambientes acuosos normales al acelerar las tasas de reacción mediante enzimas (véase, por ejemplo, la revisión Hazen et al. 2002). Para las propiedades fisicoquímicas del agua a alta presión y alta temperatura, véase Basset M-P 2003 (el agua se comporta más como un solvente orgánico apolar bajo estas condiciones). La catálisis por minerales, como los presentes en las fuentes hidrotermales de aguas profundas, mejora aún más las reacciones de moléculas orgánicas en un ambiente acuoso de este tipo (2). La degradación de moléculas orgánicas sintetizadas bajo estas condiciones de alta temperatura y alta presión puede ser evitada por minerales, al menos esto se ha demostrado para los aminoácidos (véase Hazen et al. 2002). Los ácidos grasos, como fuente de material para la formación de membranas, también podrían haber sido sintetizados en fuentes hidrotermales (Orgel 2004).
Además, también existen, simplemente tibias a moderadamente calientes, ventas hidrotermales que tienen microambientes semipermeables de dimensiones celulares (imitando una membrana lipídica), los cuales podrían retener moléculas a altas concentraciones. Esto proporcionaría una posible solución al problema de la concentración (Russell y Martin 2004, Robinson 2005). Sin embargo, aún queda por ver de qué manera tales compartimentos inmóviles habrían permitido la selección natural, lo cual es posible en el caso de vesículas de ácidos grasos mediante la competencia por el material de la membrana (Chen et al. 2004) y otros recursos, como se discute aquí.
Koonin y Martin (2005) proponen que, si los microcompartimentos minerales tenían cierta porosidad, unidades evolutivas más competitivas podrían haberse extendido sobre otras ocupando preferentemente nuevos compartimentos vacíos que deberían formarse continuamente por precipitación de sulfuro en las ventas hidrotermales. Esto habría sido un progenitor de la división celular. La alternativa que, en contraste con este modelo interesante pero aún hipotético, ya goza de un amplio apoyo experimental habría sido, por supuesto, la división de vesículas de ácidos grasos que contienen polímeros genéticos mediante fuerzas fisicoquímicas simples, véase arriba. Los autores también argumentan (cf. Martin y Russell 2003) que los microcompartimentos minerales en lugar de lípidos podrían haber servido como membrana celular incluso para un organismo tan avanzado como LUCA, ya que, en su opinión, esto permitiría la explicación más simple de por qué los descendientes de LUCA, arqueobacterias y eubacterias, tienen membranas lipídicas muy diferentes; estas habrían surgido solo más tarde.
Las fuentes hidrotermales pueden ofrecer otra solución más al problema de concentración de componentes clave en el origen de la vida. Los monómeros y oligonucleótidos de ARN pudieron haberse acumulado en gran medida en sistemas de poros hidrotermales debido a un gradiente térmico a través del poro (Baaske et al. 2007, véase también el comentario acompañante, Koonin 2007). Hasta 1000 veces la concentración de nucleótidos se logra en un solo poro, y más de 108 veces en poros concatenados (estos poros alcanzan tamaños de hasta milímetros, por lo tanto mucho más grandes que los microcompartimentos anteriores). Esta acumulación no depende de superficies de adsorción que podrían ser restrictivas para secuencias de reacciones. El grupo de Jack Szostak (Budin et al. 2009) demostró que también los ácidos grasos se concentran en gran medida en sistemas similares. Al concentrar soluciones diluidas tanto de ADN como de ácidos grasos, la autoensamblaje de grandes vesículas que contienen ADN encapsulado ocurre en regiones de los capilares donde se supera la concentración crítica de agregación del ácido graso.
Todos estos escenarios de concentración pueden funcionar no solo en fuentes hidrotermales de aguas profundas, sino también en sistemas hidrotermales que se encuentran en aguas someras en la periferia de volcanes. La convección térmica dentro de los poros hidrotermales (Baaske et al. 2007) también habría permitido el ciclo térmico para separar el producto de doble cadena de las reacciones de copia de ARN.
La química en estos entornos podría abrir nuevas posibilidades para la síntesis de un polímero genético anterior al ARN, o del propio ARN, necesario en el escenario de los genes primero.
6. Especificidad de las reacciones químicas y el escenario del metabolismo primero
Algunos extienden los hallazgos anteriores de las fuentes hidrotermales de aguas profundas a una variante del escenario del metabolismo primero, en contraposición al escenario del gen primero descrito anteriormente, una hipótesis que afirma que los ciclos metabólicos complejos podrían haberse autoorganizado, independientemente de un sistema genético capaz de proporcionar catalizadores poliméricos. Según esta hipótesis, solo a través de tal metabolismo estable la síntesis de nucleótidos y oligonucleótidos, necesarios para el inicio del mundo del ARN, habría sido posible; sin embargo, debido a los recientes hallazgos aquí discutidos, la síntesis prebiótica espontánea de ARN se está volviendo ahora mucho más probable que cuando originalmente se propusieron estos escenarios del metabolismo primero.
En las mismas publicaciones en las que Günter Wächtershäuser había predicho la síntesis de moléculas orgánicas bajo condiciones como las encontradas en los respiraderos hidrotermales, predicciones que han sido confirmadas experimentalmente (véase arriba), también introdujo un modelo metabólico (Wächtershäuser 1988, Wächtershäuser 1990). Esta hipótesis inventiva y detallada integra de manera impresionante una multitud de observaciones en el campo de la química. Sugiere que el comienzo de la vida fue lo que se ha denominado una vida plana, un metabolismo bidimensional elaborado en las superficies minerales de los respiraderos hidrotermales de aguas profundas; esto también aborda el problema de la dilución concentrando toda la química en la superficie. Central para esta y otras hipótesis de metabolismo primero es el ciclo del ácido cítrico reductor (ciclo de Krebs inverso) que proporciona un mecanismo central de biomoléculas útiles a partir de CO2 (Morowitz et al. 2000, Smith y Morowitz 2004).
Sin embargo, el desarrollo de este y otros escenarios relacionados (por ejemplo, Smith y Morowitz 2004) requeriría una sorprendente falta de reacciones secundarias (mientras que un nivel residual pequeño de reacciones secundarias podría favorecer el desarrollo evolutivo). No es fácil ver cómo la extraordinaria especificidad de las reacciones químicas, necesaria para secuencias complejas de reacciones y metabolismo para el funcionamiento de la vida, sería en general posible sin polímeros catalíticos que presenten una bolsa de sustrato tridimensional. Estos solo se proporcionan en un escenario de genes primero. Leslie Orgel (Orgel 1998) comenta sobre el asunto de la siguiente manera (para una crítica extendida, véase Orgel 2000):
No hay acuerdo sobre el grado en que el metabolismo podría desarrollarse independientemente de un material genético. En mi opinión, no hay base en la química conocida para la creencia de que largas secuencias de reacciones puedan organizarse espontáneamente y todo motivo para creer que no pueden. El problema de lograr una especificidad suficiente, ya sea en solución acuosa o en la superficie de un mineral (3), es tan severo que la probabilidad de cerrar un ciclo de reacciones tan complejo como el ciclo del ácido cítrico inverso, por ejemplo, es despreciable. Lo mismo, creo, es cierto para ciclos más simples que involucran moléculas pequeñas que podrían ser relevantes para el origen de la vida y también para ciclos basados en péptidos.
Por lo tanto, parece razonable asumir que el desarrollo de los ciclos y vías metabólicas habría requerido polímeros genéticos/catalíticos, aunque, obviamente, las opiniones están divididas sobre el asunto.
Los grandes conjuntos autocatalíticos de Stuart Kauffman (Kauffman 1993) pueden ser demasiado optimistas por la misma razón. Los conjuntos autocatalíticos tienen más probabilidades de realizarse cuando son pequeños.
Sin embargo, los desarrollos más recientes pueden parecer que plantean la posibilidad de que el escenario del metabolismo primero sea realista después de todo. Se encontró (Zhang y Martin 2006) que tres de los cinco pasos reductivos del ciclo inverso de Krebs (Fig. 3) podrían ser impulsados por partículas de ZnS (proporcionando poder reductor de electrones de la banda de conducción y creído que fueron prevalentes en las aguas de la Tierra primitiva) bajo la influencia de la luz UV. Las conversiones de oxalacetato a malato, y de fumarato a succinato, procedieron con rendimientos asombrosos (75 % y 95 %, respectivamente), que pueden estar cerca de lo suficiente. La conversión de succinato a oxoglutarato también procedió, aunque a una tasa baja del 2,5 %. Los autores sugieren que un ensamblaje mineral más complejo que solo el ZnS podría impulsar todo el conjunto de reacciones.

Figura 3. Ciclo inverso de Krebs. Se resaltan las cinco reacciones de reducción (etiquetadas 1-5). De (Zhang y Martin 2006). Reproducido con permiso. Copyright 2006 American Chemical Society.
De alto interés son también los hallazgos recientes que han sido reportados (Robinson 2005) para dar esperanza a los adherentes del modelo del metabolismo primero. Se ha demostrado (Cordova et al. 2005a) que tales moléculas orgánicas simples como aminoácidos individuales pueden catalizar la síntesis estereoespecífica de azúcares desde materiales de partida simples con especificidad similar a la de las enzimas, aunque solo en disolventes orgánicos. Por ejemplo, ya sea L- o D-enantiómeros de ciertos aminoácidos (con la serina entre ellos, véase más abajo) pueden desencadenar la formación de un cierto tipo de azúcar no solo con excelente quimioselectividad (i.e., evitando reacciones secundarias no deseadas), sino también la formación de uno de los 16 posibles enantiómeros de este azúcar con aproximadamente 99 % de estereoespecificidad. Estas reacciones se basan en el principio de la organocatálisis asimétrica descubierto en la década de 1970 (Eder et al. 1971, Hajos y Parrish 1974).
Aún más prometedoras son las conclusiones de un estudio de seguimiento (Cordova et al. 2006). En este trabajo, los autores informan que los pequeños péptidos (principalmente díptidos probados, o péptidos con no más de cinco aminoácidos) (4) y los tetrazoles de aminoácidos pueden catalizar reacciones de aldol, algunas de ellas produciendo azúcares, con gran estereoespecificidad en agua, no solo en disolventes orgánicos.
¿Podría la catálisis altamente específica por aminoácidos u otras moléculas orgánicas pequeñas (5) de manera más general, es decir, no solo en esta reacción particular, sustituir en cierta medida a las bolsas de sustrato tridimensionales de los polímeros catalíticos? ¿Podría cerrar los ciclos metabólicos?
Estas posibilidades especulativas, que involucran una catálisis específica por moléculas orgánicas, se acercarían más al escenario del metabolismo primero que fue propuesto por Christian De Duve (De Duve 1995). Él asume un protometabolismo que involucra un mundo de tioéster que también proporciona energía para las reacciones moleculares las cuales habrían llegado a ser la base para la emergencia del mundo del ARN (este escenario fue sugerido para la sopa prebiótica, no para la química en las fuentes hidrotermales). Sin embargo, De Duve basa su catálisis en multímeros derivados de tioésteres, estructuralmente similares a las [primeras] pequeñas proteínas catalíticas. Es difícil imaginar cómo tales unidades catalíticas relativamente grandes y complejas podrían haberse formado constantemente con alta reproducibilidad de manera espontánea. Tal reproducibilidad habría sido necesaria para establecer y mantener un metabolismo estable, o de otro modo, secuencias complejas de reacciones; por supuesto, no plantea ningún problema para los polímeros genéticos, como lo proporciona el escenario del gen primero. Sin embargo, la reproducibilidad (y abundancia) de la síntesis de supuestos pequeños catalizadores de moléculas sin genes probablemente habría sido un problema mucho menor.
¿Podría el único entorno acuoso de alta presión y alta temperatura de las fuentes hidrotermales de aguas profundas, que produce cambios drásticos en la reactividad de los compuestos orgánicos (véase arriba), también hacer que pequeñas moléculas orgánicas actúen como catalizadores específicos que no realizarían esta función en una solución acuosa normal?
Finalmente, para resolver el principal
enigma del modelo de genes primero: ¿podría la catálisis por pequeñas moléculas
orgánicas estar incluso involucrada en la síntesis de nucleótidos y oligonucleótidos
en la forma de los estereoisómeros correctos, una química mucho más compleja que
simplemente la síntesis de azúcares, en ausencia de polímeros genéticos/catalíticos
precursoras?
La investigación futura podría informarnos sobre todos estos temas.
Cabe señalar que la dependencia de la luz UV
para las reacciones descritas del ciclo reductor del ácido cítrico (Zhang y Martin 2006), posiblemente en conjunto con un requisito de temperaturas moderadas bajo las cuales se reportó que procedían estas reacciones, limitaría esta química a la superficie de la Tierra. Como
Leslie Orgel señala (Orgel 2006), aunque históricamente los defensores del metabolismo primero
han argumentado mayormente por la síntesis en fuentes hidrotermales de aguas profundas y
los defensores de genes primero por una sopa prebiótica en la superficie de la Tierra, ninguna
de estas conexiones puede ser un requisito a priori para el respectivo modelo:
el metabolismo sin genes podría haberse desarrollado en la superficie de la Tierra, o genes
sin metabolismo previo en fuentes hidrotermales.
Por otro lado, aunque tanto estas reacciones del ciclo del ácido cítrico reductor como la nueva síntesis de monómeros de ribonucleótidos de pirimidina activados (Powner et al. 2009) se reportan que requieren luz UV, las reacciones que aparentemente están confinadas a un solo lugar pueden demostrarse, a medida que avanza el conocimiento, que también proceden en otros lugares bajo diferentes condiciones. La síntesis de monómeros de ARN en sistemas hidrotermales podría ser un escenario atractivo ya que, como se discutió anteriormente, los poros hidrotermales permitirían una gran acumulación de monómeros y oligómeros de ARN por los gradientes térmicos a través de ellos (Baaske et al. 2007).
7. Origen de la homochiralidad de los aminoácidos y azúcares
La vida sintetiza casi exclusivamente L-aminoácidos y D-azúcares. Una pregunta clave relacionada con el origen de la vida es el surgimiento de esta homociralidad (unidireccionalidad). La homociralidad es esencial para el funcionamiento de las proteínas como polímeros de aminoácidos, y para la estructura del ADN y el ARN, que requiere la incorporación de D-azúcares.
¿Cómo surgió esta homociralidad de los aminoácidos y los azúcares? Es una pregunta que ha desconcertado a los investigadores del origen de la vida durante décadas, sin embargo, una serie de hallazgos recientes aborda el problema de manera asombrosamente bien.
Los enantiómeros (formas L- o D-) de los aminoácidos pueden enriquecerse significativamente en dos pasos:
1. Un desequilibrio inicial de las formas enantioméricas de un aminoácido
¿Cómo pudo surgir, por muy mínimo que fuera, una mayor presencia de una forma enantiomérica de un aminoácido sobre la otra (exceso enantiomérico, abreviado: ee)? Después de todo, la síntesis típica de un aminoácido en el laboratorio resulta en una relación exacta de 1:1 entre los enantiómeros L y D, una mezcla racémica.
Una posible fuente son los meteoritos. En el meteorito de Murchison, un ejemplo bien estudiado, se ha reportado que la forma L de algunos de los aminoácidos encontrados está presente hasta en un 9 % ee (Cronin y Pizzarello 1997). Estos ee pueden haber sido inducidos por luz UV polarizada circularmente (Bailey et al. 1998). Un estudio reciente (Glavin y Dworkin 2009) reporta ee de L-isovalina superiores al 18 % en el meteorito de Murchison, y postula como causa la alteración acuosa (un proceso en el que los cambios en los minerales ocurren influenciados por el agua). Aminoácidos como la isovalina, que no es común en la biosfera terrestre, pueden haber transferido quiralidad a compuestos biogénicos como catalizadores en reacciones químicas (Pizzarello y Weber 2004).
Sin embargo, si bien la evolución química de la vida primitiva pudo haberse basado muy bien en material meteorítico, las fuentes de excesos enantioméricos leves e incluso pronunciados de aminoácidos pudieron haber surgido en numerosos lugares de la Tierra prebiótica.
Robert Hazen y sus colegas encontraron (Hazen et al. 2001) que los cristales del mineral común formador de rocas calcita (CaCO3) pueden preferir adsorber las formas D- o L- del ácido aspártico (y en experimentos preliminares, las formas D- o L- de alanina también) dependiendo de la quiralidad de la superficie del cristal. El ee promedio fue de unos pocos puntos porcentuales. De manera similar, la adsorción de ácido 3-carboxi adipico, una molécula relevante prebióticamente, a cristales de calcita o feldespato produjo ee de hasta 10 % (Castro-Puyana et al. 2008).
Aunque tales leves excesos enantioméricos locales pueden ser suficientes para desencadenar eventos que puedan amplificarlos fuertemente (véase más abajo), otro estudio encontró preferencias mucho mayores de adsorción de aminoácidos a superficies minerales (Wedyan y Preston 2005). Mientras que en su estudio la cuarzo, la caolinita y la montmorillonita mostraron leves preferencias por la adsorción de enantiómeros, los sedimentos ordinarios de estuarios exhibieron una fuerte selectividad. Las relaciones típicas D/L eran enormemente diferentes de 1, alcanzando en su extremo más alto hasta 100 para la serina. Los sedimentos se incineraron para eliminar la materia orgánica que podría introducir un sesgo quiral. Los autores son cautelosos: La posibilidad de que el proceso de incineración realmente esculpa y active la superficie mineral mientras quema la materia orgánica natural (¿quiral?) no puede descartarse. Sin embargo, también señalan que es notable que ocurra tal fuerte selectividad en absoluto.
Se ha reportado un mecanismo completamente diferente y atractivo para lograr exceso enantiomérico (Kojo et al. 2004), que implica la formación de una fase sólida de cristales de aminoácidos. Esto podría haber ocurrido fácilmente en un paisaje prebiótico mediante el enfriamiento de una solución acuosa concentrada y cálida de aminoácidos o simplemente por la evaporación lenta de una solución. La mayoría de los aminoácidos racémicos forman cristales que también son racémicos. Sin embargo, cuando el asparagina D, L-racémica forma cristales, estos no son racémicos, sino que muestran grados variables de exceso de cualquiera de los enantiómeros L o D. Cuando están presentes otros aminoácidos, ya sea su forma L o su forma D se co-cristaliza preferentemente con la forma enantiomérica (L o D) del asparagina que confiere un exceso durante la formación del cristal en cuestión.
El comportamiento de los aminoácidos en el estudio mostró una gran consistencia. Si había un sesgo hacia la L-asparagina en un cristal, prácticamente todos los otros 12 aminoácidos co-cristalizados mostraron también un sesgo hacia la forma L. El grado de sesgo hacia la L-asparagina variaba entre cristales; cuando el sesgo era más pronunciado, el sesgo hacia la forma L en los otros aminoácidos también era más pronunciado. Los efectos observados podían ser muy altos. Si en otro cristal había un sesgo hacia la forma D de la asparagina, los otros aminoácidos co-cristalizados exhibieron también un sesgo hacia su enantiómero D.
Por supuesto, el equilibrio enantiomérico general de la mezcla completa de aminoácidos (la suma de todos los cristales y la fase líquida sobre ellos) seguía siendo racémica. Sin embargo, si estos tipos de cristales se formaron a partir de una solución en la Tierra prebiótica y algunos de los cristales fueron posteriormente separados físicamente de otros, lo cual podría haber ocurrido mediante muchos procesos ordinarios, y luego se redisolieron, los aminoácidos en solución habrían exhibido automáticamente un ee. Dependiendo del cristal particular del que derivaba la solución, este ee podría haber sido alto.
Otra manera de lograr exceso enantiomérico dentro de un racemato son condiciones especiales de cristalización muy lejos del equilibrio. Normalmente, el ácido aspártico y el ácido glutámico forman cristales racémicos. Sin embargo, cuando se someten a ascenso capilar a través de un material poroso (como un ladrillo parcialmente sumergido en una solución), lo cual va acompañado de sobresaturación debido a la evaporación de la solución a través de la estructura porosa, se forman cristales puros L- o D- (Viedma 2001). Condiciones similares podrían haber estado presentes en la Tierra primitiva en ambientes sedimentarios, como en una playa o una barra de arena.
2. Mejora del exceso enantiomérico mediante equilibrios de fase sólida-líquida
Los estudios han demostrado que, una vez que existe un exceso inicial de un enantiómero en una mezcla de aminoácidos, incluso si es muy leve, puede tener un efecto enorme. Este efecto puede ocurrir cuando los aminoácidos sólidos y disueltos de dicha mezcla coexisten en equilibrio, es decir, cuando los cristales se forman, por ejemplo, tras una evaporación limitada de una solución.
Un estudio detallado fue realizado por el grupo de Donna Blackmond (Klussmann et al. 2006). Cuando una mezcla de enantiómeros L- y D- de un aminoácido muestra un exceso de un enantiómero, en la mayoría de los casos un equilibrio de aminoácido sólido y disuelto consistirá en los siguientes dos o tres componentes:
a) cristales racémicos (cristales con una proporción 1:1 de los enantiómeros L- y D-, sin exceso enantiomérico en ninguno de los dos)
b) cristales puros del enantiómero en exceso
(Sea que solo esté presente una de estas dos fases sólidas, o que ambas coexistan, dependerá del exceso enantiomérico (ee) global.)
c) una solución de aminoácido en equilibrio con la(s) fase(s) sólida(s) que también exhibe un cierto ee.
Sin embargo, como resulta, para varios aminoácidos, este ee en solución es mucho mayor que el ee global de la mezcla total (el material sólido, dominado por cristales racémicos, muestra por lo tanto un ee correspondientemente menor). En el caso más extremo, la serina proporciona una solución casi enantiopura (> 99 % ee) en agua a partir de una muestra casi racémica (solo alrededor de 1 % ee) bajo condiciones de equilibrio sólido-líquido.
Un estudio más pequeño, realizado de forma independiente al mismo tiempo, reporta hallazgos similares (Breslow, Levine 2006). La evaporación lenta de una solución acuosa de fenilalanina con solo un 1 % ee del enantiómero L dio lugar a una solución de este aminoácido con un 40 % ee del enantiómero L sobre material sólido. Si, a su vez, dicha solución se dejaba evaporar, la solución resultante en equilibrio con el material sólido tenía un 90 % ee.
En un estudio más reciente, el grupo de Blackmond (Klussmann et al. 2007) amplió el concepto a mezclas de aminoácidos con otros compuestos, que pueden co-cristalizar con los aminoácidos. Demostraron que, al influir en la solubilidad, en algunos casos estos compuestos influyeron fuertemente en la ee en solución bajo condiciones de equilibrio sólido-líquido. Por ejemplo, bajo esas condiciones la ee de la valina aumentó del 47 % hasta un 99 % en presencia de ácido fumárico. Tenga en cuenta que la plausibilidad prebiótica se ve reforzada en este escenario, ya que emplea mezclas de compuestos en lugar de componentes puros.
Otra forma de aumentar la ee no emplea el enriquecimiento de la ee en solución, sino en la fase sólida de un equilibrio sólido-líquido. Un curioso mecanismo de crecimiento de cristales, que implica la abrasión por molienda, permite la aparición de un único estado quiral en la fase sólida a partir de un desequilibrio inicial pequeño en la composición del cristal tan bajo como 2-3 % ee (Noorduin et al. 2008). Sin embargo, sí requiere la racemización del compuesto en solución y se ha aplicado a un derivado de aminoácido que experimenta esta racemización tras la adición de base. Se discute una explicación teórica que difiere de la proporcionada en el estudio en un comentario sobre los hallazgos (McBride y Tully 2008). En un estudio de seguimiento, los hallazgos se extendieron a un aminoácido propiamente dicho, el ácido aspártico (Viedma et al. 2008). La racemización en solución se logró con salicilaldehído catalítico en medio ácido, y se obtuvo hasta un 99 % ee en cristales a partir de un desequilibrio inicial de menos del 10 % ee. El calentamiento podría sustituir a la molienda mecánica.
En los estudios de Klussmann et al. 2006 y de Breslow, Levine 2006, los hallazgos se discuten también en términos de catálisis por aminoácidos de reacciones aldólicas, que incluyen la síntesis de azúcares y podrían resultar en azúcares D enantiopuros; véanse también los hallazgos de Córdova et al. 2005a y Córdova et al. 2006 revisados anteriormente. Sin embargo, cuando se considera la nueva ruta de síntesis para monómeros de ribonucleótidos pirimidínicos activados (Powner et al. 2009), que evita las moléculas de base y de azúcar y su condensación, sino que produce las moléculas finales mediante intermediarios que aportan átomos tanto a la porción de azúcar como a la de base de los ribonucleótidos, la relevancia de la catálisis por aminoácidos en la producción de azúcares D o cualquier otra posible implicación de aminoácidos enriquecidos en enantiómeros en la síntesis de nucleótidos no es clara. Dado que la gliceraldehida es una de las moléculas de partida en la nueva síntesis (Fig. 1), la quiralidad en el ARN podría haber surgido por un aporte de gliceraldehida homoquiral, cuyo origen podría haber resultado de procesos similares a los discutidos para los aminoácidos.
En cualquier caso, también se ha demostrado que una vez que estaba presente un exceso enantiomérico de aminoácidos, las reacciones catalizadas por ellos podrían haber aumentado aún más el exceso enantiomérico. Se demostró que en la catálisis de la síntesis de azúcares por aminoácidos puede ocurrir una amplificación significativa del exceso enantiomérico (Cordova et al. 2005b). Por ejemplo, una reacción mediada por el aminoácido prolina con un exceso enantiomérico tan bajo como 40 % aún produjo casi azúcar hexosa enantiopuro. La reacción catalizada por prolina con un 10 % de exceso enantiomérico proporcionó al azúcar un 33 % de exceso enantiomérico.
*****
En resumen, un escenario plausible para el surgimiento de la homociralidad en la Tierra prebiótica puede concebirse a partir de todos los hallazgos descritos:
1. Origen de excesos enantioméricos locales, leves o pronunciados, de aminoácidos
2. En caso de que estos excesos enantioméricos sean solo leves, una tremenda amplificación mediante equilibrios de fase sólida-líquida; estos también pueden potenciar excesos enantioméricos ya altos
Como hemos visto en el ejemplo de la síntesis de azúcar, la catálisis por aminoácidos podría haber desempeñado un papel en la aparición de otros productos quirales relevantes para el origen de la vida, y la amplificación quiral también podría haber ocurrido durante el proceso.
[Si los enantiómeros específicos de péptidos pequeños fueron los catalizadores (cf. arriba), podrían haber surgido por conjugación de aminoácidos después del enriquecimiento enantiomérico.]
*****
Una vez que surge la homociralidad en algunas biomoléculas, puede aparentemente imponer la homociralidad correspondiente a otras. Hemos visto esto con la catálisis de aminoácidos, pero también podría aplicarse a la relación entre el ARN y los aminoácidos. Una vez que los azúcares D estuvieron presentes en el ARN, el paso hacia los aminoácidos L en las proteínas podría haber sido automático. La síntesis de proteínas requiere la aminoacilación del ARN, y es en este paso donde los aminoácidos L podrían haber sido seleccionados, como muestra un estudio elegante (Tamura, Schimmel 2004).
En el estudio, se utilizó un minihélice de ARN que recapitula el dominio dentro de los ARN de transferencia que alberga el sitio de unión de aminoácidos. Se cree que tales minihélices de ARN son los precursores de los ARN de transferencia modernos, los cuales presentan aminoácidos unidos a ellos a la maquinaria ribosomal que sintetiza proteínas. Los autores demostraron que el minihélice de ARN fue aminoacilado por aminoácidos activados con una clara preferencia por los aminoácidos L en lugar de los aminoácidos D. Un sistema de ARN de imagen especular mostró la selectividad opuesta.
Se probaron tres aminoácidos: alanina, leucina y fenilalanina, y la selectividad observada fue de 4 veces. Los autores señalan en una publicación posterior, que estudia el mecanismo (Tamura, Schimmel 2006), que un efecto de 4 veces, repetido bajo presión selectiva muchas veces, puede llevar a una preferencia abrumadora por un aminoácido L en un sistema biológico.
Por otro lado, las cadenas peptídicas también podrían haber adquirido quiralidad de manera independiente al ARN, véase por ejemplo Saghathelian et al. 2001, véase también comunicado de prensa, Hitz y Luisi 2004, Weissbuch et al. 2004, Plankensteiner et al. 2005. Por supuesto, cualquier mecanismo de este tipo en péptidos también podría haber sido ayudado por un exceso enantiomérico de aminoácidos, como se logró en los escenarios descritos anteriormente.
Cualquiera que haya sido la secuencia precisa de eventos en el origen de la vida, la fuerza acumulativa de todos los datos anteriores indica que el misterio de la quiralidad del origen de la vida ahora puede explicarse, después de todo.
*****
El tema de la quiralidad, entre otros, ha sido promocionado por los creacionistas como un "gran problema" para el concepto de un origen de la vida por causas naturales. Alegadamente, solo una intervención milagrosa de Dios podría haber resuelto el problema. Sin embargo, los hallazgos anteriores son un ejemplo típico de por qué el concepto de Dios de los huecos no funciona: la ciencia cierra rápidamente los huecos que anteriormente podrían haber sido pensados como reservados para una intervención milagrosa.
Esto es exactamente lo que debería esperarse si el mundo material es todo lo que existe, o si el mundo fue creado por un Dios que, como causa primaria, eligió crear a través de causas secundarias precisamente aquellas causas naturales que estudia la ciencia. De hecho, los creacionistas deberían preguntarse seriamente si su concepto de Dios no es uno que menosprecia: el Diseñador Inteligente como "reparador" que se ve obligado a romper sus propias leyes de la naturaleza creadas de vez en cuando porque son insuficientes para lograr ciertas etapas en el desarrollo del mundo material. Desde una perspectiva filosófica teísta, los hallazgos reales de la ciencia sugieren una idea mucho más grandiosa de Dios: el Diseñador que estableció un conjunto elegante y autosuficiente de leyes de la naturaleza que logran el despliegue de su creación induciendo la autoorganización del mundo material. Esta idea es fácilmente compatible con el concepto de Dios de muchas religiones mainstream, incluidas la mayoría de las cristianas.
Los hiperciclos, como una organización general de conjuntos autocatalíticos, han sido propuestos como un modelo para el origen de la vida (Eigen y Schuster 1977) y se explican en Principia Cybernetica Web. Sin embargo, los hiperciclos complejos solo existen como simulaciones por ordenador. Más de treinta años después de la introducción de la hipótesis, no existe ninguna evidencia experimental para hiperciclos complejos, posiblemente prebióticos. Esto los convierte aún en nada más que mera especulación y, aunque la hipótesis ha sido un tema recurrente en la literatura científica sobre el origen de la vida, ya no se menciona con frecuencia en las publicaciones recientes del campo.
Hasta ahora, solo se ha generado un hiperciclo muy simple hace más de 10 años en el laboratorio con dos péptidos autorreplicantes por el grupo de Ghadiri (Lee et al. 1997). Sin embargo, estos autores publicaron una corrección (Lee et al. 1998) en la que declaran: Aunque los datos cinéticos sugieren la intermediación de especies de orden superior en los procesos autocatalíticos, el sistema actual no debería denominarse un ejemplo de hiperciclo mínimo en ausencia de evidencia experimental directa del acoplamiento cruzado autocatalítico entre replicadores.
Existe un único informe sobre un hiper ciclo que ocurre naturalmente (Eigen et al. 1991). Pero este ejemplo, relacionado con el ciclo de infección de un bacteriófago de ARN, obviamente no es prebiótico; el ciclo evolucionó en la complejidad de los seres vivos como plantilla.
Aunque la investigación reciente y emocionante ha proporcionado escenarios plausibles para el origen de la vida y ha respondido muchas preguntas, es claro que queda mucho por investigar, ya que gran parte de los escenarios sobre el origen de la vida sigue siendo hipótesis. Se necesitan modelos experimentales que sean tanto realistas como de cierta complejidad apreciable. Por ejemplo, si fuera posible demostrar que se podría construir en el laboratorio un organismo primitivo de ARN (Szostak et al. 2001), sería un paso significativo hacia adelante. Para esto, véase el artículo de Carl Zimmer; allí también se expresa la esperanza de que la evolución de tal organismo podría ser observable en el banco de laboratorio.
(1) Fuera de un contexto celular, la evolución del ARN en el laboratorio es ahora rutinaria. La ingeniería de ARN, según mecanismos que se asemejan en cierto modo a la evolución, ha demostrado producir muchos catalizadores de ARN (véase, por ejemplo, Robertson et al. 2004). Ciertamente, todos los procedimientos sofisticados utilizados en el laboratorio no habrían estado disponibles para un precursor antiguo de una célula. Por otro lado, nada en la probeta se asemeja en absoluto a las posibilidades que surgirían de la competencia de células replicantes.
(2) La hipótesis de que la vida podría haber originado en fuentes hidrotermales resulta, en cierta medida, atractiva también debido al hecho de que los sulfuros metálicos, encontrados en esas ubicaciones y capaces de catalizar reacciones orgánicas simples, siguen encontrándose en los centros catalíticos de enzimas metabólicas centrales como la ferredoxina, la succinato deshidrogenasa y la acetil-CoA sintasa bacteriana, posiblemente sugiriendo una preservación evolutiva de la catálisis primitiva por sulfuros metálicos.
(3) Wchtershuser presenta interesantes argumentos teóricos sobre por qué el metabolismo superficial, a diferencia de las reacciones en solución, debería abordar la especificidad de reacción de manera suficiente, sin embargo, su punto de vista también ha sido criticado por Christian De Duve y Stanley Miller en un artículo conjunto (De Duve, Miller 1991). La evidencia experimental sobre la especificidad de una secuencia de reacciones estrictamente confinada a una superficie mineral (no solo catalizada por ella, lo que permitiría la difusión de productos intermedios lejos de y de vuelta a las superficies catalíticas) sería difícil de obtener, y además las propias publicaciones experimentales de Wchtershuser, así como las de otros que prueban su escenario de metabolismo primero, hasta ahora no abordan realmente este problema tan central para su hipótesis, sino que se concentran en otros aspectos más simples de su modelo.
(4) Varios ejemplos de cómo podría haber ocurrido la oligomerización prebiótica de aminoácidos a péptidos pequeños:
a) A través de la reacción de formación de péptidos inducida por sal (SIPF), posiblemente en charcos, pozas de marea o lagunas (Schwendinger, Rode 1989)
b) Por gas volcánico (Leman et al. 2004)
c) En fuentes hidrotermales de aguas profundas, mediante activación con monóxido de carbono (CO) en presencia de minerales (Huber et al. 2003)
(5) Altamente específica principalmente en términos de quimioselectividad; en términos de estereoespecificidad solo donde sea aplicable. Esta catálisis específica también podría tener lugar junto con la formación de complejos en superficies minerales, como las encontradas en fuentes hidrotermales de aguas profundas, o las moléculas orgánicas podrían formar complejos con metales disueltos en este ambiente de alta temperatura y alta presión (cf. Cody et al. 2000).
Baaske P, Weinert FM, Duhr S, Lemke KH, Russell MJ, Braun D (2007) Acumulación extrema de nucleótidos en sistemas de poros hidrotermales simulados. Proc. Natl. Acad. Sci. USA 104: 9346-9351 [Pubmed] [Texto completo gratuito]
Bada JL, Fegley B Jr, Miller SL, Lazcano A, Cleaves HJ, Hazen RM, Chalmers J (2007) Debate sobre la evidencia del origen de la vida en la Tierra. Science 315:937-939 [Pubmed]
Bailey J, Chrysostomou A, Hough JH, Gledhill TM, McCall A, Clark S, Menard F, Tamura M (1998) Polarización circular en regiones de formación estelar: implicaciones para la homociralidad de biomoléculas. Science 281: 672-674 [Pubmed]
Basset M-P (2003) ¿Es el agua a alta presión la cuna de la vida? JPCM 15: 1353-1361 [Resumen, texto completo gratuito disponible allí]
Bokov K, Steinberg SV (2009) Un modelo jerárquico para la evolución del ARN ribosómico 23S. Nature 457: 977-980 [Pubmed]
Breslow R, Levine SM (2006) Amplificación de concentraciones enantioméricas bajo condiciones prebióticas creíbles. Proc. Natl. Acad. Sci. USA 103:12979-12980 [Pubmed] [Texto completo gratuito]
Budin I, Bruckner RJ, Szostak JW (2009) Formación de vesículas similares a protocélulas en una columna de difusión térmica. J. Am. Chem. Soc. 131: 9628-9629 [Pubmed] [Artículo completo en la página de publicaciones del laboratorio de Szostak]
Castro-Puyana M, Salgado A, Hazen RM, Crego AL, Alegre ML (2008) La primera contribución de la electroforesis capilar al estudio de los orígenes abióticos de la homociralidad: Investigación de la adsorción enantioselectiva de ácido 3-carboxi-adípico en minerales. Electroforesis 29:1548-1555 [Pubmed]
Chen IA, Roberts RW, Szostak JW (2004) El surgimiento de la competencia entre protocélulas modelo. Science 305:1474-1476 [Pubmed] [Artículo completo en la página de publicaciones del laboratorio de Szostak]
Chyba C, Sagan C (1992) Producción endógena, entrega exógena y síntesis por impacto de moléculas orgánicas: un inventario para los orígenes de la vida. Nature 355:125-132 [Pubmed]
Chyba CF (2005) Reconsiderando la atmósfera temprana de la Tierra. Science 308: 962-963 [Pubmed]
Cleaves HJ, Chalmers JH, Lazcano A, Miller SL, Bada JL (2008) Una reevaluación de la síntesis orgánica prebiótica en atmósferas planetarias neutras. Orig. Life Evol. Biosph. 38:105-115 [Pubmed]
Cody GD, Boctor NZ, Filley TR, Hazen RM, Scott JH, Sharma A, Yoder HS Jr. (2000) Compuestos de hierro-azufre carbonilados primordiales y la síntesis de piruvato. Science 289:1337-1340 [Pubmed]
Cordova A, Ibrahem I, Casas J, Sunden H, Engquist M, Reyes E (2005a) Neogénesis de carbohidratos catalizado por aminoácidos: una transformación antigua plausible. Chemistry 11: 4772-4784 [Pubmed]
Cordova A, Engquist M, Ibrahem I, Casas J, Sunden H. (2005b) Orígenes plausibles de la homociralidad en la neogénesis de carbohidratos catalizada por aminoácidos. Chem. Commun. (Camb) (2005): 2047-2049 [Pubmed]
Cordova A, Zou W, Dziedzic P, Ibrahem I, Reyes E, Xu Y (2006) Reacciones aldol intermoleculares asimétricas directas catalizadas por aminoácidos y pequeños péptidos. Chemistry 12: 5383-5397 [Pubmed]
Cronin JR, Pizzarello S (1997) Excesos enantioméricos en aminoácidos meteoríticos. Science 275: 951-955 [Pubmed]
De Duve C, Miller SL (1991) ¿Vida bidimensional? Proc. Natl. Acad. Sci. USA 88: 10014-10017 [Pubmed] [Texto completo gratuito]
De Duve, C (1995) Los comienzos de la vida en la Tierra. American Scientist 83: 428-437 [Texto completo gratuito]
Eder U, Sauer G, Wiechert R (1971) Nuevo Tipo de Ciclización Asimétrica para Esteroideos Ópticamente Activos CD Estructuras Parciales. Angew. Chem. Int. Ed. Engl. 10:496-497
Eigen M, Schuster P (1977) El hiperciclo. Un principio de autoorganización natural. Parte A: Emergencia del hiperciclo. Naturwissenschaften 64: 541-565
Eigen M, Biebricher CK, Gebinoga M, Gardiner WC (1991) El hiperciclo. Acoplamiento de la biosíntesis de ARN y proteínas en el ciclo de infección de un bacteriófago de ARN. Biochemistry 30:11005-11008
Gil R, Silva FJ, Pereto J, Moya A (2004) Determinación del núcleo de un conjunto mínimo de genes bacterianos. Microbiol. Mol. Biol. Rev. 68: 518-537 [Pubmed] [Texto completo gratuito]
Glass JI, Assad-Garcia N, Alperovich N, Yooseph S, Lewis MR, Maruf M, Hutchison CA 3rd, Smith HO, Venter JC (2006) Genes esenciales de un bacterio mínimo. Proc. Natl. Acad. Sci. USA 103: 425-430 [Pubmed] [Texto completo gratuito]
Glavin DP, Dworkin JP (2009) Enriquecimiento del aminoácido L-isovalina por alteración acuosa en los cuerpos padres de los meteoritos CI y CM. Proc. Natl. Acad. Sci. USA 106: 5487-5492 [Pubmed]
Hajos ZG, Parrish DR (1974) Síntesis asimétrica de intermediarios bicíclicos de la química de productos naturales. J. Org. Chem. 39:1615-1621 [Resumen]
Hanczyc, MM, Fujikawa, SM, Szostak, JW (2003) Modelos experimentales de compartimentos celulares primitivos: Encapsulación, crecimiento y división. Science 302:618-622 [Pubmed] [Artículo completo en la página de publicaciones del laboratorio de Szostak]
Hanczyc MM, Szostak JW (2004) Vesículas replicantes como modelos del crecimiento y división de células primitivas. Curr. Opin.Chem. Biol. 8:660-664 [Pubmed] [Artículo completo en la página de publicaciones del laboratorio de Szostak]
Hanczyc MM, Mansy SS, Szostak JW (2007) Ensamble de membranas dirigido por superficies minerales. Orig. Life Evol. Biosph. 37:67-82 [Pubmed] [Artículo completo en la página de publicaciones del laboratorio de Szostak]
Hazen, RM, Filley TR, Goodfriend GA (2001) Adsorción selectiva de aminoácidos L- y D sobre calcita: implicaciones para la homociralidad bioquímica. Proc. Natl. Acad. Sci. USA 98:5487-5490 [Pubmed] [Texto completo gratuito]
Hazen RM, Boctor N, Brandes JA, Cody GD, Hemley RJ, Sharma A, Yoder Jr HS (2002) Alta presión y el origen de la vida. JPCM 14: 11489-11494 [Resumen]
Hazen RM (2005) Gen-e-sis: La Búsqueda Científica del Origen de la Vida, Joseph Henry Press
Hitz TH, Luisi PL (2004) Inicio espontáneo de la homociralidad en cadenas de oligopéptidos generadas en la polimerización de aminoácidos de N-carboxianhídrido en agua. Orig. Life Evol. Biosph. 34:93-110 [Pubmed]
Huang W, Ferris JP (2006) Síntesis de un solo paso y regioselectiva de hasta 50-meros de oligómeros de ARN mediante catálisis de montmorillonita. J. Am. Chem. Soc. 128: 8914-8919 [Pubmed]
Huber C, Wächtershäuser G (1997) Ácido acético activado por la fijación de carbono sobre (Fe,Ni)S bajo condiciones primitivas. Science 276:245-247 [Pubmed]
Huber C, Eisenreich W, Hecht S, Wchtershuser G (2003) Un posible ciclo de péptidos primordiales. Science 301: 938-940 [Pubmed]
Huber C, Wächtershäuser G (2006) Ácidos alfa-hidroxil y alfa-amino bajo posibles condiciones de origen de la vida hadeanas y volcánicas. Science 314:630-632 [Pubmed]
Johnson AP, Cleaves HJ, Dworkin JP, Glavin DP, Laczano A, Bada JL (2008) El experimento de descarga eléctrica volcánica de Miller. Science 322:404 [Pubmed]
Johnston WK, Unrau PJ, Lawrence MS, Glasner ME, Bartel DP (2001) Polimerización de ARN catalizada por ARN: extensión de cebador templada por ARN precisa y general. Science 292: 1319-1325 [Pubmed]
Joyce GF (2002) La antigüedad de la evolución basada en ARN. Nature 418:214-221 [Pubmed]
Kauffman, Stuart. (1993) Orígenes del orden: autoorganización y selección en la evolución, Oxford University Press [Amazon]
Kasting JF, Zahnle KJ, Walker JCG (1983) Fotoquímica del metano en la atmósfera primitiva de la Tierra. Precambrian Res. 20: 121-148 [Resumen]
Klussmann M, Iwamura H, Mathew SP, Wells DH Jr, Pandya U, Armstrong A, Blackmond DG (2006) Control termodinámico de la amplificación asimétrica en la catálisis de aminoácidos. Nature 441: 621-623 [Pubmed]
Klussmann M, Toshiko I, White AJP, Armstrong A, Blackmond DG (2007) Emergencia de la homociralidad en fase de solución mediante ingeniería de cristales de aminoácidos. J. Am. Chem. Soc. 129: 7657-7660 [Pubmed]
Kojo S, Uchino H, Yoshimura M, Tanaka K (2004) La asparagina racémica D,L causa un exceso enantiomérico de otros aminoácidos racémicos D,L coexistentes durante la recristalización: una hipótesis que explica el origen de los aminoácidos L en la biosfera. Chem. Commun. (2004): 2146-2147 [Pubmed]
Koonin EV (2007) Un reactor para la síntesis de ARN en el origen de la vida. Proc. Natl. Acad. Sci. USA 104: 9105-9106 [Texto completo gratuito]
Koonin EV, Novozhilov AS (2009) Origen y evolución del código genético: el enigma universal. IUBMB Life 61:99-111 [Pubmed]
Koonin EV, Martin W (2005) Sobre el origen de los genomas y las células dentro de compartimentos inorgánicos. Trends Genet. 21:647-654 [Pubmed]
Kuhn WR, Atreya SK (1979) Fotólisis de amoníaco y el efecto invernadero en la atmósfera primitiva de la Tierra. Icarus 37: 207-213 [Pubmed]
Kun A, Santos M, Szathmary E (2005) Ribozimas reales sugieren un umbral de error relajado. Nat. Genet. 37:1008-1011 [Pubmed]
Lee DH Severin K, Yokobayashi Y, Ghadiri MR (1997) Emergencia de la simbiosis en la autorreplicación de péptidos a través de una red hipercíclica. Nature 390:591-594 [Pubmed]
Lee DH Severin K, Yokobayashi Y, Ghadiri MR (1998) Erratum: Emergencia de la simbiosis en la autorreplicación de péptidos a través de una red hipercíclica Nature 394:101
Leman L, Orgel LE, Ghadiri MR (2004) CFormación prebiótica de péptidos mediada por sulfuro de carbonilo. Science 306: 283-286 [Pubmed]
McBride JM, Tully JC (2008) Química física: ¿Se detuvo la vida antes de comenzar? Nature 452:161-162 [Pubmed]
Mansy SS, Schrum JP, Krishnamurthy M, Tob S, Treco DA, Szostak JW (2008) Síntesis dirigida por plantilla de un polímero genético en un protocelula modelo. Nature 454:122-125 [Pubmed] [Artículo completo en la página de publicaciones del laboratorio de Szostak]
Mansy SS, Szostak JW (2008) Estabilidad térmica de membranas de protocélulas modelo. Proc. Natl. Acad. Sci. USA 105: 13351-13355 [Pubmed] [Texto completo gratuito]
Martin W, Russell MJ (2003) Sobre el origen de las células: una hipótesis para las transiciones evolutivas desde la geoquímica abiótica hasta los procariotas quemoautótrofos, y desde los procariotas hasta las células nucleadas. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 358:59-83 [Pubmed] [Texto Completo Gratuito]
Miller SL (1953) Producción de aminoácidos bajo posibles condiciones primitivas de la Tierra. Science 117 (1953): 528-529 [PDF]
Morowitz HJ, Kostelnik JD, Yang J, Cody GD (2000) El origen del metabolismo intermedio. Proc. Natl. Acad. Sci. USA 97: 7704-7708 [Pubmed] [Texto completo gratuito]
Noorduin WL, Izumi T, Millemaggi A, Leeman M, Meekes H, Van Enckevort WJ, Kellogg RM, Kaptein B, Vlieg E, Blackmond DG (2008) Surgimiento de un único estado sólido quiral a partir de un derivado de aminoácido casi racémico. J. Am. Chem. Soc. 130: 1158-1159
Orgel LE (1998) El origen de la vida: una revisión de hechos y especulaciones. Trends Biochem. Sci. 23: 491-495 [Pubmed]
Orgel LE (2000) Ciclos bioquímicos autoorganizadores. Proc. Natl. Acad. Sci. USA 97: 12503-12507 [Pubmed] [Texto completo gratuito]
Orgel LE (2004) Química prebiótica y el origen del mundo del ARN. Crit. Rev. Biochem. Mol. Biol. 39: 99-123 [Pubmed]
Orgel LE (2006) Síntesis Geotérmica y Metabolismo. Astrobiology 6: 297-298
Pavlov AA, Kasting JF, Brown LL, Rages KA, Freedman R (2000) Calentamiento por efecto invernadero por CH4 en la atmósfera de la Tierra primitiva. J. Geophys. Res. 105: 11981-11990 [Pubmed]
Pizzarello S, Weber AL (2004) Aminoácidos prebióticos como catalizadores asimétricos. Science 303:1151
Plankensteiner K, Reiner H, Rode BM (2005) Diferenciación estereoselectiva en la reacción de formación de péptidos inducida por sal y su relevancia para el origen de la vida. Péptidos 26:535-541 [Pubmed]
Poole AM (2006) Obtener una transición desde un mundo de ARN hacia células modernas se ha vuelto un poco más fácil. BioEssays 28:105-108 [Pubmed]
Powner MW, Gerland B, Sutherland JD (2009) Síntesis de pirimidinas activadas en condiciones prebiológicamente plausibles. Nature 459: 239-242 [Pubmed]
Rajamani S, Vlassov A, Benner S, Coombs A, Olasagasti F, Deamer D (2008) Síntesis de polímeros similares al ARN a partir de mononucleótidos asistida por lípidos. Orig. Life Evol. Biosph. 38:57-74 [Pubmed]
Rees
M (2001) Solo seis números: las fuerzas profundas que dan forma al universo,
Basic Books [Amazon]
Ricardo A, Carrigan MA, Olcott AN, Benner SA (2004)
Los minerales de borato estabilizan la ribosa. Science 303:196
Robertson MP, Knudsen SM, Ellington AD (2004) Selección in vitro de ribozimas dependientes de péptidos para la actividad. RNA 10:114127 [Pubmed] [Texto completo gratuito]
Robinson R (2005) Arrancando un mundo celular: Investigando el origen de la vida, desde sopa hasta redes. PLoS
Biol 3(11): e396 [Texto completo gratuito]
Russell MJ, Martin W (2004) Las raíces rocosas de la ruta del acetil-CoA. Trends Biochem. Sci. 29: 358-363 [PDF]
Sagan C, Chyba C (1997) La paradoja del sol temprano débil: el escudo orgánico de los gases de efecto invernadero lábiles ante los ultravioletas. Science 276:1217-1221[Pubmed]
Saghatelian A, Yokabayashi Y, Soltani K, Ghadiri MR (2001) Un replicador peptídico quiroselectivo. Nature 409:797-801 [Pubmed]
Schaefer L, Fegley Jr. B (2007) Desgasificación de material condrita ordinaria y algunas de sus implicaciones para la química de asteroides, planetas y satélites. Icarus 186:462-483 [Resumen] [PDF]
Schwendinger MG, Rode BM (1989) Posible papel del cobre y el cloruro de sodio en la evolución prebiótica de péptidos. Analytical Science 5: 411-444 [Resumen] [Texto completo gratuito]
Smith E, Morowitz HJ (2004) Universalidad en el metabolismo intermedio. Proc. Natl. Acad. Sci. USA 101: 13168-13173 [Pubmed] [Texto completo gratuito]
Smolin L (1999) La vida del cosmos, Oxford University Press [Amazon]
Susskind L (2006) The Cosmic Landscape: String Theory and the Illusion of Intelligent Design, Back Bay Books [Amazon]
Szostak JW, Bartel DP, Luisi PL (2001) Sintetizando la vida. Nature 409: 387-390 [Artículo completo en la página de publicaciones del laboratorio de Szostak]
Szostak JW (2009) Química de sistemas en la Tierra primitiva. Nature 459: 171-172
Tamura K, Schimmel P (2004) Aminoacilación selectiva por quiralidad de un minihélice de ARN. Science 305:1253 [Pubmed]
Tamura K, Schimmel P (2006) Aminoacilación selectiva por quiralidad de un minihélice de ARN: características mecanísticas y supresión quiral. Proc. Natl. Acad. Sci. USA 103:13750-13752 [Pubmed] [Texto completo gratuito]
El Mundo del ARN. (2006) Editado por: Gesteland RF, Czech TR, Atkins JF. Cold Spring Harbor Laboratory Press [Pubmed]
Tian F, Toon OB, Pavlov AA, De Sterck H (2005) Una atmósfera temprana de la Tierra rica en hidrógeno. Science 308: 1014-1017 [Pubmed]
Viedma C (2001) Cristalización enantiomérica a partir de DL-aspártico y DL-glutámico: implicaciones para la quiralidad biomolecular en el origen de la vida. Orig. Life Evol. Biosph. 31: 501-509 [Pubmed]
Viedma C, Ortiz JE, de Torres T, Izumi T, Blackmond DG (2008) Evolución de la homociralidad de fase sólida para un aminoácido proteogénico. J. Am. Chem. Soc. 130:15274-15275 [Pubmed]
Wächtershäuser G (1988) Antes de las enzimas y las plantillas: teoría del metabolismo superficial. Microbiol. Rev. 52: 452-484 [Texto completo gratuito]
Wächtershäuser G (1990) Evolución de los primeros ciclos metabólicos. Proc. Natl. Acad. Sci. USA 87: 200-204 [Pubmed] [Texto completo gratuito]
Walker JC, Brimblecombe P (1985) Hierro y azufre en el océano prebiológico. Precambrian
Res. 28:205-222 [Pubmed]
Wedyan M, Preston MR (2005) Adsorción selectiva de isómeros de aminoácidos por componentes de sedimentos naturales. Environ. Sci.
Technol. 39:2115-2119 [Pubmed]
Weissbuch I, Bolbach G, Leiserowitz L, Lahav M (2004) Amplificación quiral de oligopéptidos mediante polimerización en cristalitas bidimensionales sobre agua. Orig. Life Evol. Biosph. 34:79-92 [Pubmed]
Wolf YI, Koonin EV (2007) Sobre el origen del sistema de traducción y del código genético en el mundo del ARN mediante selección natural, exaptación y subfuncionalización. Biol. Direct 2:14 [Pubmed] [Texto completo gratuito]
Zaher HS, Unrau PJ (2007) Selección de un ribozima de ARN polimerasa mejorado con
extensión y fidelidad superiores. RNA 13: 1017-1026 [Pubmed] [Texto completo gratuito]
Zahnle KJ (1986) Fotoquímica del metano y la formación de ácido cianhídrico (HCN) en la atmósfera primitiva de la Tierra. J. Geophys. Res. 91:
2819-2834 [Resumen]
Zhang L, Peritz A, Meggers E (2005) Un ácido nucleico glicólico simple. J. Am.
Chem. Soc. 127:41744175 [PubMed]
Zhang XV, Martin ST (2006) Impulsar partes del ciclo de Krebs en reversa mediante fotoquímica mineral. J. Am. Chem. Soc.
128: 16032-16033 [PubMed]
Zhu TF, Szostak JW (2009) Crecimiento acoplado y división de membranas de protocélulas modelo. J. Am. Chem. Soc. 131: 5705-5713 [PubMed] [Artículo completo en la página de publicaciones del laboratorio de Szostak]
[Volver a las preguntas frecuentes sobre abiogénesis]