O Sistema Complementar é Irredutivelmente Complexo?
por Mike Coon![]()
"Por complexidade irredutível, eu me refiro a um único sistema que é composto por várias partes interagentes que contribuem para a função básica, e onde a remoção de qualquer uma das partes faz com que o sistema cesse efetivamente de funcionar. Um sistema irredutivelmente complexo não pode ser produzido gradualmente por modificações sucessivas e sutis de um sistema precursor, já que qualquer precursor de um sistema irredutivelmente complexo é, por definição, não funcional."
-- Michael Behe
Este ensaio tem como objetivo reforçar alguns elementos da excelente FAQ de Keith Robison, que detalha as falhas nos argumentos de Behe (http://www.talkorigins.org/faqs/behe/review.html). Outra excelente refutação por H. Allen Orr pode ser encontrada em http://bostonreview.net/br21.6/orr.html. Veja também http://bostonreview.net/evolution.html para vários artigos sobre design inteligente e complexidade irredutível por Orr, Russell F. Doolittle, Douglas J. Futuyma, Richard Dawkins, Michael Behe, Phillip Johnson e David Berlinski, entre outros.
Muitas pessoas no Talk.Origins (T.O.) e em outros lugares apresentaram as razões conceituais pelas quais a evolução darwiniana pode produzir sistemas complexos irredutivelmente (IC). Aqui, tento apresentar algumas evidências para apoiar esses conceitos. No FAQ de Robison, ele mencionou que não sabia se o sistema complemento exibia um sistema de cascata variável evolutivamente. Tal variabilidade ao longo de linhas filogenéticas ilustraria a falácia central por trás do conceito de complexidade irredutível de Behe. Essa falácia decorre da incapacidade de perceber que o que agora parece ser complexidade irredutível é simplesmente o resultado da evolução darwiniana, na qual características que outrora eram apenas úteis agora se tornam essenciais. Abaixo, tento abordar o ponto de Robison e mostrar como o sistema complemento falha no teste de Behe.
A propósito, um ponto que Robison fez em seu FAQ foi em refutação à alegação de Behe de que os anticorpos não podem funcionar por conta própria e, portanto, são IC. Behe está errado pelo menos na medida em que os anticorpos possuem funções além do papel que desempenham na cascata do complemento. Há até algumas evidências de que os anticorpos possuem uma função fora de seu papel convencional de "marca molecular". Não entrarei em detalhes aqui, mas um bom exemplo seria o fato de que os anticorpos podem ser catalíticos. Ou seja, podem funcionar não apenas como marcas moleculares em patógenos, tumores ou células infectadas por vírus, mas eles podem ter funções enzimáticas que são bastante independentes de seus papel imunológicos principais (veja, por exemplo, Barbas1).
Primeiro, uma breve revisão do sistema complemento2
Em 1899, o belga Jules Bordet descobriu que o soro continha dois componentes: um termoestável que apresentava atividade de aglutinação e precipitação, posteriormente identificado por Paul Ehrlich como anticorpos, e um termolábil responsável pela lise bacteriana. No entanto, não foi até às décadas de 1950 que foi elucidado o primeiro sistema de proteínas do soro termolábeis (mais de 30 foram identificadas desde então), cuja função consistia em complementar a ação antibacteriana dos anticorpos. Ainda mais cedo, embora mal compreendido, o papel das proteínas do soro termolábeis foi notado primeiramente em tentativas iniciais de transfusão sanguínea, e estas observações podem ajudar-nos a compreender como o complemento funciona.
A maioria das pessoas sabe que uma das partes mais importantes do nosso sistema imunológico é um tipo de proteína chamado anticorpo. Uma célula imunológica especializada chamada célula B produz anticorpos. As células B são linfócitos que desempenham papéis-chave na erradicação de infecções. Primariamente, elas se desenvolvem em um tipo de célula conhecido como Célula Plasmática, que produz grandes quantidades de anticorpos, embora também desempenhem um papel como células apresentadoras de antígenos e no recrutamento de células T. Nossos anticorpos são mais frequentemente produzidos para reconhecer patógenos invasores e, portanto, não são prejudiciais para nós. Mas às vezes os anticorpos podem causar danos e os clínicos se referem a uma maneira para isso acontecer como reação hemolítica de transfusão, que é frequentemente fatal. O cenário mais comum, raro nos dias de hoje, é a transfusão de sangue do tipo A em um indivíduo do tipo O. O receptor do tipo O produz anticorpos contra os glóbulos vermelhos do doador do tipo A. Logo após a transfusão, o receptor desenvolve calafrios, febre, urticária e sangramento descontrolado. O plasma do receptor fica rosa, devido à vazamento de hemoglobina dos glóbulos vermelhos do doador. O sangue é esgotado de fatores de coagulação, porque o sistema de coagulação foi ativado e coágulos sanguíneos se formam difusamente dentro da circulação.
No entanto, descobriu-se que a ligação do anticorpo às células vermelhas do tipo A não as danifica. Se misturarmos células vermelhas suspensas em soro fisiológico com anticorpo puro, as células aglomeram-se, mas não vazam hemoglobina. Quando se adiciona plasma sanguíneo às células aglomeradas, as células lisam, ou seja, estouram, e se administrarmos o plasma de volta a um receptor (num modelo animal), replica as outras características de uma reação hemolítica de transfusão. Claramente, algo no plasma está a realizar estes efeitos, não o anticorpo. Esta atividade foi denominada "complemento" porque complementa a ação do anticorpo. Agora sabemos muito sobre a composição e a função desta importante função imunitária e identificámos três formas pelas quais o complemento pode eliminar patógenos invasores.
Proteínas do complemento ativadas iniciam uma cascata de reações enzimáticas que resulta, em última instância, em uma grande abertura literalmente perfurada na membrana do microrganismo invasor, ou, na reação hemolítica de transfusão, nos glóbulos vermelhos. Nossas próprias células são protegidas das vias do complemento ativadas espontaneamente (veja abaixo) porque possuem outras proteínas que inativam a cascata do complemento. Não surpreendentemente, alguns microrganismos evoluíram uma estratégia de defesa que envolve a presença de seus próprios inibidores de cascata. Mas a maioria não possui essa defesa e o complexo proteico que perfura o microrganismo, chamado perforina, cria um grande poro na membrana do patógeno. Essa perda de integridade da membrana destrói a capacidade do patógeno de controlar a concentração de sais dentro de si, matando-o assim.
Nossas funções efetoras complementares atuam como braços tanto do sistema imune adaptativo quanto do sistema imune inato. O sistema imune inato está presente em todos os organismos multicelulares, enquanto a imunidade específica (ou adaptativa) é encontrada apenas nos vertebrados, com exceção dos agnatos, ou peixes sem mandíbula. Os receptores de antígenos no sistema inato são codificados pelo genoma germinativo e reconhecem amplamente estruturas homólogas (geralmente essenciais) compartilhadas por grupos de microrganismos. O sistema imune adaptativo, no entanto, desenvolve seu repertório de receptores de antígenos por meio de mutações somáticas, e muitos dos determinantes reconhecidos por esses receptores de antígenos são idiossincráticos (geralmente não essenciais e, portanto, mutáveis) para um microrganismo particular. O sistema imune inato, ao contrário da imunidade específica, não desenvolve memória. Ambos os sistemas têm a capacidade de distinguir o próprio, mas o sistema imune específico é imperfeito nesse aspecto, resultando em autoimunidade.
Os três caminhos complementares são armas que um organismo pode utilizar para ajudar a combater patógenos, destruir tumores e eliminar células infectadas por vírus. São conhecidos como os caminhos lectina, alternativa e clássica (existem algumas variações menores em alguns organismos). O caminho da lectina, às vezes chamado de caminho lítico, e o caminho alternativo fazem parte da resposta imune inata e, portanto, são considerados evolutivamente mais antigos do que o caminho clássico (veja abaixo). O caminho da lectina é iniciado por receptores do complemento compostos por lectinas séricas circulantes (as lectinas são proteínas que se ligam a grupos de açúcar), que se ligam a moléculas de superfície de patógenos contendo resíduos de manose (por exemplo, manano), enquanto o caminho alternativo é iniciado pelo complemento que se liga à superfície de um patógeno e é ativado espontaneamente. O terceiro caminho, conhecido como caminho clássico por razões históricas (foi descoberto primeiro), é ativado pela opsonização por anticorpos de patógenos (ligação de anticorpos a epítopos específicos de proteínas de superfície, por exemplo, antígenos) e, portanto, faz parte da imunidade adaptativa.
Cada um dos três caminhos possui eventos iniciais distintos, incluindo a ligação de receptores e um conjunto único de componentes do sistema complemento (veja abaixo) que iniciam uma cascata enzimática. Todos os três caminhos convergem em uma atividade enzimática chamada C3 convertase. Para vertebrados, tanto no caminho lectina quanto no clássico, a enzima C3 convertase é uma proteína conhecida como C2b, enquanto no caminho alternativo a enzima convertase é uma combinação funcional e estruturalmente homóloga de duas proteínas, C3b e o fator B plasmático. Esta combinação, por sua vez, é ativada pelo fator protease plasmático D. Neste ponto, todos os três caminhos possuem funções efetoras semelhantes. Abaixo segue um esquema dos principais atores em todos os três caminhos de ativação do complemento em vertebrados* (por favor, note: infelizmente, muitas das proteínas foram numeradas na ordem em que foram descobertas, não na sua ordem respectiva nos caminhos).
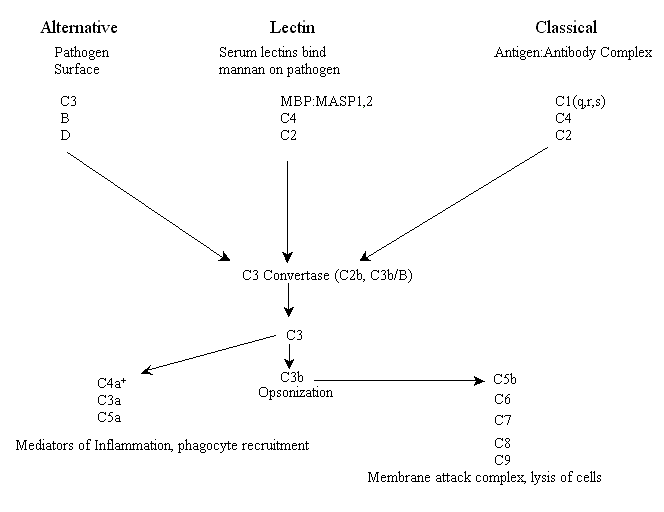
O C4a é um produto da clivagem do C4 pela MASP ou C1s, e não pelo C3, e atua como um mediador da inflamação.
|
Resumo das proteínas envolvidas na cascata do complemento |
|
|
Ligação ao complexo Ag:Ab |
C1q |
|
Ativação de enzimas |
C1r, C1s, C2b, Bd, D, MASP1,2 |
|
Opsoninas de ligação à membrana |
C4b, C3b, MBP |
|
Mediadores da inflamação |
C5a, C3a, C4a |
|
Ataque à membrana |
C5b, C6, C7, C8, C9 |
|
Receptores do complemento |
CR1, CR2, CR3, CR4, C1qR |
|
Proteínas reguladoras do complemento |
C1INH, C4bp, CR1, MCP, DAF, H, I, P, CD59 |
*Adaptado de Janeway & Travers Immunobiologia, 1996; Current Biology Ltd/Garland Publishing Inc.
História evolutiva das vias complementares
Todos os vertebrados que foram estudados, com exceção dos agnatos, compartilham os sistemas complementares humanos. À medida que nos afastamos filogeneticamente, tanto o sistema imunológico quanto seus efetores, incluindo os sistemas complementares, são diferentes. Três que foram estudados são a lampreia (um agnato), o grupo ascídia dos urocordados e os ouriços-do-mar. Houve muita discussão sobre a história evolutiva das vias complementares. Há algumas evidências de que, nos vertebrados, a via alternativa antecede a via clássica, com base em comparações de sequências do primeiro componente do complemento, C1q3. No entanto, outras evidências sugerem que a via alternativa é mais antiga que a via clássica com base em comparações filogenéticas de C3a, C3b e C5a4. Além disso, o sistema complementar da lampreia consiste apenas nas vias alternativa e lectina5. Mais recentemente, demonstrou-se que um urocordado possui genes que correspondem aos genes C3, C4 e C5 dos vertebrados, mas não ao C26. Além disso, Ji et al7 e Nonaka, M et al8 mostraram que a via lectina é evolutivamente a mais antiga das três, demonstrando que a MASP é encontrada em um urocordado, a ascídia Halocynthia roretzi (ouríngua japonesa). Acredita-se que grande parte do sistema complementar vertebrado surgiu da duplicação de genes que codificam as moléculas C3/C4/C5, Bf/C2, C1s/C1r/MASP-1/MASP-2 e C6/C7/C8/C99
O caminho complementar é complexidade irredutível?
Uma das alegações de Behe é que cascatas enzimáticas como a ativação do complemento são complexas irredutivelmente porque cada novo passo na cascata "exigiria tanto uma proenzima quanto uma enzima ativadora para ligar a proenzima no momento e local corretos". Ele aponta para o sistema do complemento, entre outros, como sendo complexos irredutivelmente. E parece mesmo. Por exemplo, todas as três cascatas dependem da atividade da C3 convertase para ativar o complemento e ajudar o organismo a se defender de patógenos. Humanos que carecem de C3 não possuem vias do complemento funcionais e sofrem com maior suscetibilidade a infecções piogênicas por bactérias patogênicas como Haemophilus influenzae e Streptococcus pneumonae, entre outras. Parece, então, que as vias dos vertebrados não poderiam ter evoluído de uma maneira darwiniana a partir de vias lectínicas anteriores, porque elas dependem da atividade da C3 convertase. Mas isso é porque Behe e outros não percebem que os atuais participantes da cascata não estavam sempre presentes.
Existem duas linhas gerais de evidência que sugerem que o sistema complemento evoluiu para o seu estado atual nos mamíferos. Uma é que observamos sistemas de complemento em invertebrados que carecem de um braço inteiro do sistema mamífero. Outra é que as proteínas dos sistemas de urocordados e vertebrados são homólogas. Em particular, existem homologias entre proteases que sugerem duplicação gênica e subsequente especialização, como notado acima. A situação é semelhante à cascata de coagulação, onde são observadas homologias que podem ser explicadas por duplicação gênica. Organismos que estão filogeneticamente distantes dos humanos e, portanto, representam uma linhagem de animais que é evolutivamente mais antiga que os vertebrados atuais, contudo, compartilham conosco a via das lectinas. Esta é uma de muitas, muitas peças de evidência para a descendência comum. Outra é o fato de que os vertebrados possuem uma via inteira que está ausente em representantes de organismos evolutivamente mais antigos.
Vamos examinar mais de perto a via da lectina, a via do complemento compartilhada por urocordados e vertebrados. Ela é iniciada pela ligação da proteína ligadora de manana (MBP) e da serina protease associada à MBP (MASP), que é encontrada como uma proenzima. A MBP reconhece resíduos de N-acetilglicosamina em proteínas de superfície de patógenos, ativando assim a MASP. A MASP ativada então cliva proteoliticamente C4 e C2. O C2 é clivado para formar C2b e C2a. O C2b também é conhecido como C3 convertase, que possui atividade enzimática comum a todas as três vias. A C3 convertase é criticamente importante para as vias de complemento em mamíferos, de modo que mutações no C2 (e no C3) podem produzir estados patológicos em ratos, camundongos, porcos-da-índia, primatas e humanos (revisado em 10-12). A deleção do gene C3 abole as vias de complemento, como observado em camundongos knockout15.
O interessante sobre a proteína C3 do tunicado marinho é que ela não contém o sítio de clivagem da convertase C3, encontrado em todas as sequências de C3 de vertebratos até à data7. Isto significa que uma enzima que é dissimilar à convertase C3 de vertebratos (C2b) ativa a proteína C3 de H. roretzi. De facto, os urocordados não têm um gene C26. Um candidato para a convertase C3 dos urocordados é o MASP. O MASP parece estar relacionado, por homologia de sequência, com as enzimas C1 da via clássica, uma das quais, a C1s, cliva o C413, 14, sugerindo assim uma origem para os genes C1 dos vertebratos através de duplicações gênicas. Existem duas enzimas MASP conhecidas por funcionarem como enzimas ativadoras nas vias do complemento. O MASP-1 liga-se à MBP (nota: a MBP é por vezes referida como MBL, por Lectina de Ligação a Manano). Esta ligação ativa o MASP-1, que então cliva o C4 e o C2, como notado acima. Foi recentemente postulado que o MASP-1 também ativa uma segunda protease de serina, chamada MASP-2, que produz uma enzima convertase C3 de vertebrato na via lectina13. Uma terceira enzima MASP, o MASP-3, foi identificada e pensa-se que regula a atividade das outras MASPs15.
O MASP em si pode ativar o C3 funcionando como uma convertase do C3? Sim. O MASP mamífero pode ativar o C3 diretamente, mas faz isso de forma deficiente16, 17 e os mamíferos incapazes de converter o C3 sofrem de uma variedade de doenças10-12. O MASP dos ascídios, no entanto, é conhecido por ter atividade de convertase do C318. Como os urocordados carecem de C2 e, portanto, de C2b, mas possuem MASP, eles parecem ter um sistema complemento truncado, mas obviamente eficaz, que contorna a necessidade da convertase do C3 C2b. Assim, embora as vias clássicas dos vertebrados sejam complexas de forma irredutível, na medida em que toda a via depende de pelo menos uma enzima, é claro que organismos evolutivamente mais antigos possuem uma via funcional mesmo na ausência dessa enzima. Na via lectina dos vertebrados, a convertase do C3 C2b assumiu a função do MASP invertebrado, de modo que agora o sistema complemento dos vertebrados depende criticamente da atividade do C2b.
Isso sugere que, à medida que os vertebrados surgiram e desenvolveram seus próprios sistemas complemento, aproveitaram a cascata dos invertebrados, mas adicionaram um componente próprio, C2 (entre outros). Isso permitiu que os vertebrados aproveitassem plenamente seu novo tipo de imunidade, a imunidade adaptativa. Eles mantiveram a via lítica, mas também estabeleceram a via complemento clássica ao cooptar o modelo invertebrado. Com o tempo, no entanto, a via clássica dos vertebrados tornou-se criticamente dependente do novo componente, C2, que as vias anteriores não exigiam. Os vertebrados também desenvolveram a via alternativa, novamente cooptando a via invertebrada.
Isso merece ser repetido: os urocordados possuem um sistema complemento funcional, mas carecem de um componente da cascata, a C3 convertase, que é essencial na mesma cascata nos vertebrados. Lembre-se da citação do Dr. Behe no início deste ensaio: "Um sistema irreducivelmente complexo não pode ser produzido gradualmente por modificações sucessivas e sutis de um sistema precursor, já que qualquer precursor de um sistema irreducivelmente complexo é, por definição, não funcional". Este exemplo de uma cascata do complemento irreducivelmente complexa que é claramente o resultado da evolução darwiniana contradiz a alegação do Dr. Behe.
Em resumo, existem problemas com a alegação de Behe de que o complemento é um "sistema complexo irredutível" ou de que é improvável que tenha surgido por evolução. Embora seja altamente articulado, a remoção de algumas partes certamente não faz com que o sistema "deixe de funcionar". A ocorrência de sistemas mais simples em organismos filogeneticamente mais antigos sugere uma via evolutiva e a ocorrência de numerosas homologias entre os genes sugere um mecanismo para sua evolução. Isso demonstra o ponto que Keith Robison, H. Allen Orr, Russell Doolittle, Douglas Futuyma e uma variedade de regulares do T.O. tentaram fazer inúmeras vezes. Para parafrasear o Dr. Orr (IIRC): "A evolução darwiniana pode facilmente produzir complexidade irredutível: tudo o que é necessário é que partes que uma vez foram apenas favoráveis se tornem, devido a mudanças posteriores, essenciais."19
Muitos agradecimentos a George Acton por suas sugestões esclarecidas e sua crítica a um rascunho deste ensaio.
Referências
3. Dodd, A.W., & F. Petry, Behring Inst Mitt 1993 (93):87-102.
4. Farries, TC, Steuer KL e JP Atkinson, Immunol Today 1990 (3):78-80.
6. Smith, L.C., Chang,, L., Britten, R.J., & E.H. Davidson, J., Immunol. 1996 (156):593-602.
7. Ji, X., Azumi, K., Sasaki, M.,& N. Masaru, Proc. Natl. Acad. Sci USA, 1997 (94):6340-6345.
9. Zarkadis IK, Mastellos D, Lambris JD, Dev Comp Immunol 2001 Out-Dez;25(8-9):745-62
10. Sakamoto M, Fujisawa Y, Nishioka K Nutrition 1998 (4):391-8.
11. Carroll MC Annu Rev Immunol 1998 (16):545-68
12. Frank M.M., J Clin Immunol 1995 (6 Suppl):113S-121S.
13. Vorup-Jensen T, Jensenius JC, Thiel S. Immunobiologia 1998 ago;199(2):348-57
14. Matsushita M, Endo Y, Fujita T. Immunobiology 1998 Aug;199(2):340-7
16. Ogata, R.T., Low, P. J. & M. Kawakami, J. Immunol. 1995 (154):2351-2357
17. Matsushita, M. & T. Fujita Immunobiologia 1995 (194):443-448
18. Nonaka M, Azumi K. Dev Comp Immunol 1999 Jun-Jul;23(4-5):421-7
19. Ver http://bostonreview.net/br21.6/orr.html.
[Retornar às Perguntas Frequentes sobre Complexidade Irredutível]