Introducción
El Dr. Lee Spetner es un teórico de la información que ha escrito un libro que afirma que las mutaciones aleatorias no pueden producir el tipo de cambios "informativos" en la biología que se alega que requiere la evolución (1). Es interesante que en un libro supuestamente sobre teoría de la información, la formulación clásica de la teoría de la información de Shannon y Weaver (2) no sea mencionada. La noción de Spetner sobre la información en la biología ha sido adoptada por varios grupos de negacionistas de la evolución, y aunque otros han producido críticas específicas de su trabajo (3,4), no existe un análisis general global de sus argumentos.
En esta revisión consideraré si las métricas de Spetner pueden aplicarse válidamente a la biología, y cómo Spetner las aplica realmente a ejemplos del mundo real. Aunque sus argumentos son superficialmente plausibles, una mirada más cercana con algún conocimiento de bioquímica revela fallos significativos. Primero describiré brevemente la métrica de información de Spetner; luego mostraré que 1) las métricas de Spetner dependen de un mecanismo de unión que no ocurre en la naturaleza, 2) que las métricas de Spetner requieren que las sustancias se unan a las enzimas de manera todo o nada, mientras que los sustratos reales no se unen de esta manera. Además, mostraré que Spetner mismo es inconsistente en la aplicación de sus métricas. En su ejemplo de Xilitol, no utiliza realmente la medida que desarrolla, y en el ejemplo de estreptomicina cambia a una métrica diferente, cuando su métrica original mostraría un aumento de información. Finalmente, mostraré que su modelo de "evolución dirigida" se basa en un malentendido de una forma de mutación aleatoria.
- Resumen de las métricas de Spetner
- Aplicación a sistemas biológicos reales:
- Ejemplos de Spetner
- Spetner y la mutación "dirigida"
- Conclusiones
- Referencias
Glosario |
| sitio activo: un
sitio de unión de un ligando (generalmente un pliegue o
hendidura en la estructura de una enzima) donde tienen lugar las reacciones
químicas. aminoácido: una pequeña molécula orgánica utilizada como bloque constructivo para péptidos o proteínas. ångström: Una distancia muy pequeña; 10-10 metro. enzima: una proteína catalítica que realiza la modificación química de un sustrato que se une a su sitio activo. ligando: cualquier molécula que se une a un sitio de unión específico en una enzima o receptor. péptido: una molécula orgánica compuesta por cadenas de aminoácidos. Las proteínas son péptidos largos. proteína: una gran molécula orgánica compuesta por cadenas de aminoácidos. receptor: una proteína que se une ligandos que son hormonas o neurotransmisores. La unión de la hormona resulta en la activación de una vía enzimática, pero la hormona en sí no se modifica químicamente (a diferencia de un sustrato). sustrato: un ligando molecular que se une a un enzima activo sitio y es modificado químicamente por la enzima. |
Resumen de las métricas de Spetner
Teorías de la información clásicas como la teoría de la información de Shannon han sido aplicadas a la biología por varios autores. Aquellos que han abordado la evolución han concluido que la evolución puede, bajo circunstancias apropiadas, aumentar la "información" en el sentido de Shannon (por ejemplo, véase 13). El Dr. Spetner, sin embargo, afirma que las mutaciones aleatorias no pueden aumentar la "información". En su afirmación, Spetner utiliza dos métricas separadas de información. Una es una medida de "expectación" mediante la cual un conjunto de cadenas diferentes tiene menos información que un conjunto de cadenas idénticas. Esto sorprenderá a las personas familiarizadas con la información estándar de Shannon Weaver o la información algorítmica, pero es una formulación válida bajo circunstancias particulares. No entraré en esto en profundidad porque Spetner tampoco lo hace.
Además, utiliza una medida de información de "direccionamiento". Esto es bastante fácil de entender.
"Brisbane" tiene menos información que
"Cannon Hill, Brisbane", que tiene menos información que
"Richmond Road, Cannon Hill, Brisbane", que tiene menos información que
"666, Richmond Road, Cannon Hill, Brisbane"
Existe una forma formal de hacerlo utilizando direcciones binarias para los elementos de matrices de n x n (o n x n x n). Es un poco forzado mapear esto sobre la unión al sustrato, pero no hay nada inherentemente más absurdo en esto que mapear la transmisión de la telegrafía a la replicación del ADN. Una característica clave de esta medida es que la "información" está directamente relacionada con la longitud de la cadena. Es decir, una cadena más larga tiene más información tanto en la formulación de la "palabra-enzima" (véase más abajo) como en la de direccionamiento binario. Otra característica importante es que la unión es "todo o nada". Discuto las implicaciones de esto en detalle en el ejemplo de la "palabra-enzima" de Spetner.
Unión enzima-sustrato y la especificidad de Spetner
Las enzimas son catalizadores proteicos que promueven reacciones químicas en los compuestos que se unen a ellas (sus sustratos). Dado que las enzimas se unen fuertemente solo a muy pocos químicos entre la multitud presente en su entorno, se consideran altamente específicas. Spetner afirma que la especificidad enzimática es análoga a la especificidad de la dirección, y que las enzimas con muchos sustratos tienen menos información que las enzimas con un solo sustrato. En su explicación utiliza un experimento mental de "enzima palabra" en lugar de un ejemplo real, y examinaremos este argumento en detalle más tarde. Sin embargo, a primera vista parece un argumento razonable. La unión enzima-sustrato (o hormona-receptor) se compara con un mecanismo de "llave y cerradura", donde el sustrato es la llave y la enzima es la cerradura. (importante advertencia: tanto la cerradura como la llave son "flexibles" ya que las enzimas y sus sustratos son algo flexibles).
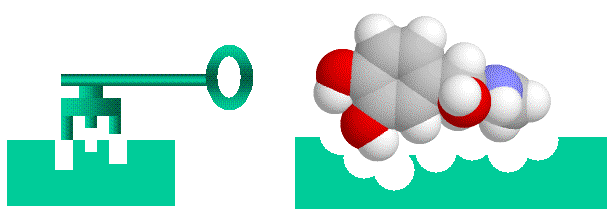 |
La especificidad de unión de Spetner no es solo una propiedad de las enzimas
Observe que el argumento de Spetner es general y se aplica no solo a las enzimas, sino a cualquier sistema de unión específico, como las interacciones hormona/receptor, la unión entre elementos del citoesqueleto, etc., independientemente de si el sustrato es actuado enzimáticamente. En la discusión siguiente utilizaré el término "ligando", que abarca todo lo que se une específicamente a un sitio aceptor, lo cual incluye los sustratos.
Observe además que hay un cambio sutil en el argumento. Es fácil ver que la dirección para "666, Richmond Road, Cannon Hill, Brisbane" requiere más información para especificar que "Brisbane", pero la afirmación de que las enzimas tienen más información porque se unen a un solo sustrato es similar a la afirmación de que "666, Richmond Road, Cannon Hill, Brisbane" tiene más información que "Brisbane" porque se entregan menos cartas allí que a "Brisbane". Esta es una afirmación algo diferente, a saber, que la información del objeto físico en una dirección tiene la información necesaria para especificar esa dirección.
Los requisitos de Spetner no se encuentran en sistemas del mundo real
Pero aceptar la afirmación de Spetner, podría parecer intuitivamente obvio que una cerradura más específica que solo acepta una llave tiene más información que una cerradura menos específica que acepta muchas llaves. Esto es en realidad incorrecto, pero en lugar de entrar en esto aquí, vea este documento sobre criptografía y llaves maestras : ADVERTENCIA archivo de 5 Mb. Por el momento aceptaremos esto. Análogamente, podríamos esperar que una enzima sea más específica porque tiene más puntos de unión que una enzima que es menos específica (y también existe la suposición no declarada de que las enzimas de un solo sustrato son más importantes que las enzimas de múltiples sustratos. No lo son).
Por ejemplo, podríamos imaginar que un sitio activo que utiliza 4 puntos de unión tiene más información que uno con dos puntos de unión
X--O O--Y X--0 O--Y
\ / \ /
DRUG DRUG
/ \
Z--N N--Y
Donde X, Y y Z son aminoácidos en el sitio activo de las enzimas.
Desafortunadamente, la biología no funciona de esa manera. El número de puntos de unión es importante, en cierto grado. Pero otras propiedades físicas de la enzima también son importantes. Por ejemplo, considera las enzimas betalactamasas que descomponen el antibiótico cefalosporinas. Una variante mutante puede descomponer cefalosporinas extendidas (donde la molécula se ha hecho más voluminosa), mientras que la enzima normal no puede. La variante mutante no tiene menos puntos de unión que la enzima normal, sino que tiene un bisagra más flexible para que el grupo catalítico pueda alcanzar el anillo lactámico con mayor facilidad.
Peor aún, puedes tener sustratos que se unen en exactamente el mismo número de puntos, pero que se unen con más firmeza porque has sustituido un átomo de cloro por un átomo de oxígeno y has cambiado la distribución de electrones en la molécula del fármaco.
La especificidad de unión no es algo simplemente análogo a la "dirección", implica forma física (que es susceptible a un análisis de dirección) y propiedades fisicoquímicas (que no son simplemente susceptibles a un análisis de dirección, especialmente cuando las propiedades son las del sustrato) y propiedades estructurales de la enzima, como la flexibilidad de la bisagra o la distorsión de la hélice, que tampoco son susceptibles a un análisis de dirección (¿una hélice que se flexiona hacia la derecha tiene más información que una hélice que se flexiona hacia la izquierda?)
En términos simples, cualquier análisis de la "información" de una proteína que aborde la especificidad de unión al sustrato, aunque superficialmente plausible, es extremadamente ingenuo, porque la especificidad del sustrato no es susceptible al análisis propuesto (especialmente cuando parte de esa información está en el sustrato). Además, su análisis se ocupa únicamente de enzimas de un solo sustrato; no sé cómo aborda las enzimas de dos sustratos y dos productos aleatorias de tipo bi-bi; parece sentir que todas las enzimas deberían ser como (algunas de) las enzimas del ciclo del ácido cítrico de Krebs.
Habiendo delineado en general por qué la métrica de Spetner no es aplicable a la unión proteína-ligando, en la siguiente sección examinaré a fondo el ejemplo de la palabra enzima de Spetner y lo compararé con un sistema receptor-ligando bien estudiado, el receptor de angiotensina II.
Aplicación a sistemas biológicos reales: [Inicio]
Ejemplo de Spetner del palabra-enzima
Examinemos esto con cierto detalle y revisemos su ejemplo de la "enzima palabra". Este difiere en aspectos clave del ejemplo de la dirección binaria que presenta primero, pero lo ignoraré en este artículo, ya que la "enzima palabra" es el ejemplo que utiliza para cerrar la brecha entre los sistemas reales [estreptomicina en su ejemplo] y la medida de dirección binaria, véase (1) páginas 134-137.
La información de Spetner es proporcional a la longitud de la cadena de unión
Aquí está la palabra enzima de Spetner "ghts". Esto puede "unirse" a muchos "sustratos" (en realidad ligandos, ya que se trata de un argumento de unión general):
ghts Nights Lights Fights Rights (and many more, omitted for clarity) Lightship Nightshade
al aumentar la longitud de la cadena, reducimos el número de "ligandos" "unidos", aumentando la "especificidad"
ghtsh Lightship Nightshade
al aumentar nuevamente la longitud de la cadena, reducimos nuevamente el número de "ligandos" "unidos", aumentando la "especificidad" una vez más.
ghtshi Lightship
En cada etapa, para cada "aumento en la especificidad", la longitud en bits de la cadena de unión aumenta. Por lo tanto, la "información" en la cadena de unión aumenta.
Observe que, aunque Spetner ha afirmado que las mutaciones aleatorias no pueden producir aumentos de información, en su ejemplo de la "enzima de palabras", las mutaciones aleatorias sí pueden aumentar la información. Simplemente añadir una letra al azar a la cadena "ghts" dará como resultado un número de cadenas que se unen a sustancialmente menos "ligandos". También dará como resultado un número de cadenas que no se unen a ningún ligando, pero es suficiente notar que la mutación aleatoria simple producirá "enzimas de palabras" de mayor "especificidad"
Sin embargo, cuando llegamos a sistemas reales, el vínculo entre el aumento de la longitud de la "cuerda" de unión y la "especificidad" simplemente no se mantiene. Un ejemplo que he dado anteriormente, un cambio en una región de bisagra, lejos del sitio de unión, puede permitir la entrada de ligandos más grandes, aunque la secuencia de unión real no cambie.
Skip text graphic a--d--------| a--d-------| a--d--------| abcd n VS abcd N abcdE N bc----------| bc---------| bc----------|
En la sección que sigue examinaré esto con mayor profundidad.
La información de Spetner depende de la unión "todo o nada"
Como se mencionó anteriormente, la información de Spetner es proporcional al número de sustratos, pero Spetner nunca define sustrato. Esto no es trivial. Como hemos visto anteriormente, el "enzima" de Spetner o bien se une a un ligando, o no lo hace. Esto es importante porque todo el núcleo del argumento de Spetner es que el número de sustratos refleja la longitud de la cadena de unión, y la longitud de la cadena de unión, en bits, es la información de la enzima/receptor/proteína de unión. Sin embargo, en el mundo real, la unión de un ligando a una proteína no es un asunto de todo o nada; los sustratos tienen grados variables de "pegajosidad", lo cual no es abordado por la métrica de Spetner. Incluso una enzima muy específica se unirá a un ligando muy fuertemente, a unos pocos no tan fuertemente, y a una gran cantidad muy débilmente. No se puede simplemente decir que solo se considerarán los ligandos muy fuertemente unidos, ya que los ligandos muy débiles pueden ser de gran importancia fisiológica si están presentes en una concentración lo suficientemente alta (hay muchos ejemplos de esto en fisiología; en el ejemplo del receptor de angiotensina II a continuación, hay un grupo de péptidos llamados péptidos de la tríada de histidina, los cuales, aunque son unidos muy débilmente al receptor de angiotensina II, efectivamente modulan la actividad del receptor bajo circunstancias fisiológicas debido a que sus concentraciones son tan altas en el cuerpo). Esto tiene una implicación teórica y práctica.
Teóricamente, esto significa que no existe una conexión necesaria entre la longitud de la cadena y el número de ligandos unidos, ya que la "pegajosidad" depende de factores moleculares que no se traducen fácilmente en información. ¿Tiene una cadena que contiene ácido aspártico más o menos información que una que contiene asparagina, cuando la diferencia virtualmente única entre ellas es la carga? La "pegajosidad" también depende de algunas características moleculares que forman parte del sustrato. Prácticamente, significa que la información de Spetner no puede medirse experimentalmente, ya que nunca podremos conocer verdaderamente el número total de sustratos a menos que probemos todos los sustratos potenciales en el universo.
Por lo tanto, podemos ver que las métricas de Spetner probablemente no sean capaces de medir el contenido de información de enzimas/receptores/proteínas de unión en absoluto. En la siguiente sección, presento un análisis exhaustivo que refuerza este punto.
Word-enzimas comparadas con proteínas reales [Inicio]
Tomemos como ejemplo los importantes receptores hormonales, los receptores de angiotensina II (AT). Los receptores AT existen en dos versiones, codificados por genes separados: el receptor AT1 es una proteína compuesta por una cadena de 359 aminoácidos, y el receptor AT2 es una proteína de 363 aminoácidos de longitud. Ambos se unen al péptido angiotensina II (AII) y a péptidos relacionados, pero la unión del AII a los receptores estimula sistemas enzimáticos completamente diferentes (la unión del AII al receptor AT1 activa la enzima fosfolipasa C, mientras que la unión al AT2 activa una enzima diferente, la fosfatasa). A pesar de tener perfiles de unión casi idénticos, los receptores comparten solo el 34% de los aminoácidos en su estructura. Tenemos una comprensión muy buena de la estructura y las propiedades del sitio de unión del AII en los receptores de angiotensina, por lo que proporcionan una plataforma para probar las ideas de Spetner sobre la especificidad.
Los ligandos AII y similares se unen a la misma secuencia de aminoácidos en los receptores AT1 y AT2. Podemos utilizar el código de un solo letra para aminoácidos, donde una sola letra sustituye al nombre de un aminoácido (por ejemplo, D es el aminoácido aspártico, véase más abajo), para representar la secuencia de unión como una cadena, análoga a la cadena de unión de palabras de Spetner ghtshi. La cadena de unión del receptor de angiotensina es DNKH. Por lo tanto, estos receptores parecen ser un buen ejemplo para probar las ideas de Spetner. Los aminoácidos en la cadena de unión no son contiguos, sino que están separados por muchos aminoácidos (5, 6).
Los aminoácidos que componen la secuencia DNKH:
D(Aspartato 281) N(Asparagina 111) K(Lisina 199) H(Histidina 256) AT1
D(Aspartato 297) N(Asparagina 126) K(Lisina 215) H(Histidina 273)
AT2
Observe que los aminoácidos se encuentran en posiciones diferentes en los distintos receptores (por ejemplo, el Aspártico 281 está en la posición 281 en la cadena de 359 aminoácidos que es el receptor AT1 y el Aspártico 297 está en la posición 297 en la cadena de 363 aminoácidos que es el receptor AT2) en parte porque el receptor AT2 tiene un C-terminal más largo, y también debido a una inserción en la secuencia entre N y K y entre K y H en el receptor AT2. Por lo tanto, el concepto de "secuencia precisa" no es exactamente aplicable a la unión de receptores. Los aminoácidos se acercan (pero no en una secuencia lineal, más bien como un anillo) mediante el plegamiento tridimensional de las proteínas, y este plegamiento tridimensional se produce gracias a secuencias muy diferentes.
|
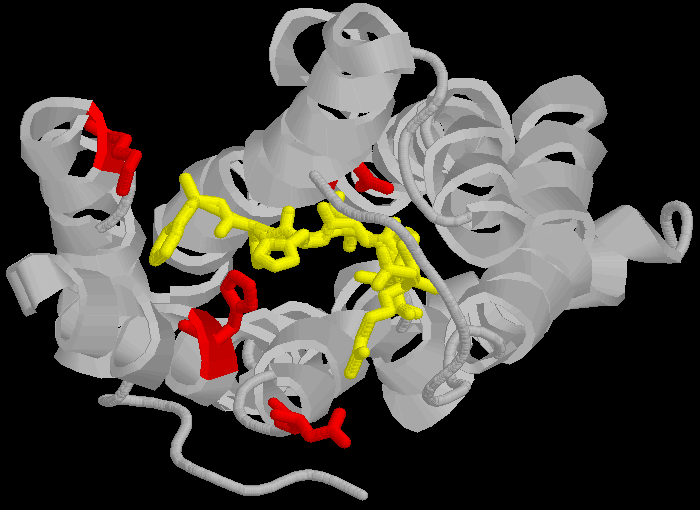
Esto es problemático para las afirmaciones de Spetner, ya que la secuencia DNKH depende de una serie de factores que NO dependen de dicha secuencia, por lo tanto, el "verdadero" contenido de información no se refleja únicamente por la secuencia de unión. De nuevo, se debe notar que el plegamiento tridimensional no es rígido y cambia entre un número de diferentes estados (las cadenas de proteínas son algo flexibles, más como una bola de espaguetis semicocidos que las formas rígidas en las que a veces se las dibuja, por lo tanto, las enzimas y receptores son candados "blandos" y los ligandos son llaves "blandas": esto es relevante para las afirmaciones de Spetner, más adelante).
Como indiqué anteriormente, los receptores AT unen la angiotensina y péptidos similares al sitio de unión del ligando. Al igual que con el receptor, no toda la secuencia del ligando se une al sitio de unión. Las secuencias se muestran en el código de un solo letra de aminoácidos, con los aminoácidos que se unen mostrados en negrita.
AII DRVYIHPF AIII RVYIHPF SarIle SRVYIHPI SarAsp SDVYIHPI CGP NYKRHPI AIV VYIHPF
Para AII y AIII, los péptidos se unen a la secuencia del sitio de unión del receptor de la siguiente manera (5,6,7,8):
RYF
DNKH
Análogo en términos generales al "enlace" de ghtshi-lightship. F se une tanto a K como a H. Esto se debe principalmente a interacciones electrostáticas en lugar de los "abultamientos y huecos" físicos que Spetner utiliza en su (incorrecta) analogía para el enlace de la estreptomicina. El aminoácido básico R se une electrostáticamente al aminoácido ácido D, Y y N forman enlaces de hidrógeno y F forma un enlace pi con H, y un enlace de hidrógeno con Y. (En SarAsp, el ácido D interactúa con el ácido D, pero la distribución de carga dipolar significa que se unen en lugar de repelerse. La química y el sentido común NO van bien juntos. (5,6))
Pero al observar SarIle y SarAsp, podemos ver que el modelo de unión de Spetner va muy mal. Estos solo tienen dos de los tres puntos de coincidencia presentes en AII y AIII, sin embargo, se unen muy bien de hecho. Quizás solo la secuencia de unión de DN sea crítica. AIV, que carece de la región de unión de D, aún se une (no tan bien como los demás, pero aún se une, más sobre esto más tarde (5,6,7,8)).
En términos del modelo de Spetner, es como si "ghtshi" uniera Lights, Nights, y Ship, así como lightship. Esto rompe el nexo que Spetner ha intentado establecer entre la longitud de la cadena de unión y el número de ligandos unidos.
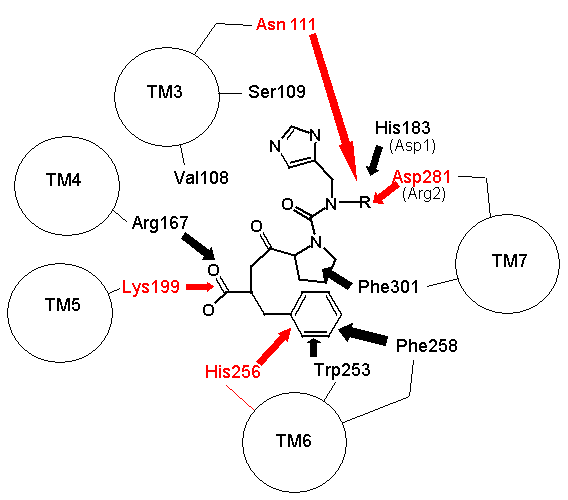  |
Peor aún, el CGP se une a aminoácidos completamente diferentes en la hendidura de unión del ligando, por lo que existe una desconexión adicional con el modelo de Spetner, ya que hay más de una "cadena de unión" en un sitio de unión de ligando. Este no es un fenómeno inusual, y también es cierto para muchos receptores. La Figura 2 muestra la comparación entre los puntos de unión de AII y losartan en el receptor AT1, mostrando las diferencias.
El número de ligandos unidos no está simplemente relacionado con la longitud de la cadena de unión
Entonces, ¿qué sucede cuando acortamos la cadena de unión DNKH. Según el modelo de Spetner, deberías aumentar el número de ligandos unidos. Podemos mutar el aminoácido H a A (alanina), Q (glutamina) o R (arginina). La mutación Q mantiene la carga y el tamaño, pero pierde el enlace pi; la mutación R mantiene la carga; la mutación A pierde la carga y el tamaño; por lo tanto, las mutaciones de H a R y A son como truncar la cadena de unión a DNK. En el receptor AT1, la unión no cambia con ninguna de estas mutaciones. Cuando mutamos H a Q o R en el receptor AT2, se pierde todo el enlace (7,8). De cualquier manera, el modelo de Spetner está completamente equivocado sobre lo que sucede cuando se acorta la "longitud de la cadena" de una cadena de unión de ligandos (y esto nuevamente invalida su medida).
Incluso peor, ¿qué sucede cuando mutas N en DNKH (Asparagina en la posición 111 en el receptor AT1 y en la posición 126 en el receptor AT2)? Si mutas N a G (glicina), un aminoácido neutro muy corto, a diferencia de la asparagina cargada más larga, no hay contacto con el ligando ni con el G, por lo que la cadena de sitios de unión se convierte en DKH. A pesar de esta truncación, aún se unen AII, AIII, etc., pero AIV se ha vuelto más "pegajoso" (algo que Spetner no aborda realmente en su métrica). Como mencioné antes, los receptores AT se mueven entre varias formas conformacionales. La mutación N->G restringe el receptor a menos formas conformacionales, haciéndolo MÁS específico. Resulta que AII y AIII se unen a todas las formas conformacionales entre las que se mueve la enzima, pero AIV solo se une a una de las formas, aquella estabilizada por la mutación N111G (5,6). Por lo tanto, la unión es una función crítica de la forma tridimensional, no cubierta por la métrica de Spetner, y una sola mutación puede aumentar la especificidad (según los criterios de Spetner, una proteína que tiene una forma conformacional tiene más especificidad que una con múltiples formas conformacionales).
¿Qué pasaría si tomamos la métrica de Spetner al pie de la letra, mirando SOLAMENTE el número de ligandos unidos. ¿Existen otros ejemplos de mutaciones que puedan aumentar la "especificidad" en los receptores AT? Sí. En el receptor AT2, reemplazar el aminoácido Y (tirosina) en la posición 215 de la cadena AT2 con R elimina la unión de CGP, dejando intacta la unión de AII y SarIle (tenga en cuenta que Y NO forma parte de la cadena de unión DNKH). Por el contrario, la mutación de Y215 a Q elimina la unión de AII y SarIle, mientras deja intacta la unión de CGP (aquí "eliminar" significa que no hay unión significativa cuando está presente 1 mM de ligando). Así podemos ver que las mutaciones individuales pueden aumentar la "especificidad", como en el número de ligandos unidos. Así, incluso con la métrica inválida de Spetner, aún podemos demostrar que la mutación aleatoria aumenta la "especificidad".
Hemos visto que en ejemplos del mundo real, los requisitos de Spetner para que el número de sustratos sea un indicador de la información de las proteínas no se cumplen, y la métrica de Spetner es inválida. Ahora nos turnaremos a los ejemplos que Spetner presentó en su libro, y mostraremos cómo sus propios análisis de estos ejemplos fallan.
Ejemplos de Spetner [Inicio]
Ahora veamos cómo Spetner aplica su propia métrica.
Metabolismo del xilitol
Dada la extensa discusión que Spetner dedica a la especificidad de unión, y sus experimentos mentales que enfatizan que la especificidad es equivalente al número de sustratos unidos, puede resultar sorprendente que en el análisis de Spetner sobre la mutación de ribulosa a xilitol no considere realmente la especificidad de unión.
Él sí considera una medida bioquímica llamada especificidad, pero no se trata de la medida de especificidad de Spetner. Esta es una relación entre la eficiencia catalítica y la especificidad de unión. Bien, ahora dices que esto es bastante trivial, podemos trabajar con esto. Bueno, no. La eficiencia catalítica es algo completamente diferente. Definitivamente no es susceptible a una medida de "direccionamiento", y se basa en una serie de propiedades fisicoquímicas, como la distribución de carga en el sustrato, la rotación libre del grupo lateral catalítico (¿tiene un grupo catalítico que rota 10 angstroms más o menos información que uno que rota 15 angstroms?) y así sucesivamente. Críticamente, dos sustratos pueden tener la misma especificidad de unión, pero diferente eficacia catalítica, lo que confunde el análisis de la información.
Peor aún, el argumento de Spetner está relacionado con el número de sustratos sobre los que actúa (véase arriba). Sin embargo, en la mutación de la ribulosa deshidrogenasa a xilitol deshidrogenasa, en ambos casos la enzima unió 3 sustratos; según su propia definición, no ha ocurrido ningún cambio de información. La velocidad de deshidrogenación ha cambiado, pero el número de sustratos sobre los que actúa no cambia. Dos puntos deben ser enfatizados:
a) Como se señaló en la sección anterior, la métrica de Spetner es una medida que indica que una enzima que une el sustrato A y el sustrato B tiene menos información que una enzima que une solo el sustrato A (recuerde ghtsh, que "une" Lightship y Nightshade tiene menos información que ghtshi, que solo une Lightship, porque ghtshi es una cadena más larga). Esto NO funciona cuando comparamos una enzima que une 20 moléculas de sustrato A por cada 80 moléculas de sustrato B, con una enzima que une 80 moléculas de sustrato A por cada 20 moléculas de sustrato B. La longitud de la cadena de unión no necesita haber cambiado, ni haber cambiado en la dirección que exige Spetner, por lo que la medida de longitud en bits no sirve en absoluto.
b) La magnitud relativa del cambio puede deberse enteramente a cambios en la eficiencia catalítica, que no tiene contenido de información en el esquema de Spetner (ni puedo ver ninguna manera significativa de calcular la "información" en esto con cualquier métrica de información existente).
Lo que es aún peor. Spetner comparó las "especificidades" de 3 sustratos, porque eran los únicos sustratos medidos en el experimento. En realidad, la ribulosa deshidrogenasa (y la xilitol deshidrogenasa) se une a muchos más sustratos que solo esos 3 (aunque la eficiencia catalítica es muy baja para la mayoría de los sustratos unidos). Como se señaló anteriormente, sin evaluar el panel completo de sustratos, cualquier afirmación sobre especificidad es sin sentido. (Pero, ¿cuáles sustratos? si nos restringimos a los sustratos naturales, excluimos el xilitol, un azúcar sintético no encontrado en el entorno natural, pero desarrollar actividad de xilitol deshidrogenasa era el punto entero del ejercicio) si incluimos sustratos sintéticos, tenemos un potencialmente número infinito de sustratos para probar. ¿Cómo sabemos qué está realmente pasando?
La situación sigue empeorando. Spetner asumió que el punto alcanzado en el experimento (donde Ribulosa y Xilitol se descomponían a tasas aproximadamente similares por la enzima mutante) era el mejor posible, que no era posible ninguna mejora adicional. En el experimento que examinó, el punto principal era ver si se podía desarrollar la actividad sobre el xilitol, no obtener la actividad óptima. De hecho, en otros experimentos se produjeron enzimas mutantes que descomponían el xilitol 20 veces más rápido que la ribulosa (lo cual, digamos que, destruye su tesis; véase 9).
Por lo tanto, el ejemplo de la ribulosa no respalda la tesis de Spetner. Además, tenemos literalmente cientos de enzimas donde la mutación aleatoria da como resultado una alta especificidad de sustrato (ha habido una enorme cantidad de trabajo en el desarrollo de actividades enzimáticas nuevas y específicas a partir de proteínas barril alfa-beta generalistas, por ejemplo, véase 10, y recientemente se ha informado sobre la evolución de actividades de unión específicas a partir de péptidos aleatorios 11).
Unión de estreptomicina: [Inicio]
  |
Su otro ejemplo es el caso de la resistencia al antibiótico estreptomicina. La estreptomicina mata bacterias interfiriendo con el ensamblaje de proteínas en el ribosoma. La mutación del gen rspL, que codifica para la subunidad S12 de la partícula ribosomal 30S en bacterias, puede resultar en resistencia a la estreptomicina. La partícula ribosomal 30S es una estructura de múltiples subunidades que, a su vez, forma parte de la partícula ribosomal sintetizadora de proteínas. La subunidad S12 junto con el ARN 16S forma parte del centro de corrección de pruebas del sitio de unión de aceptación del ARN de transferencia (ARNt). La mutación de la lisina de unión a estreptomicina en la posición 42 de la cadena de proteínas que constituye la subunidad S12 a treonina o asparagina resulta en que la estreptomicina no se una a S12, con la consiguiente resistencia de las bacterias al antibiótico estreptomicina. Este es un ejemplo clásico de una mutación beneficiosa, como se encuentra en muchos libros de texto, ya que fue el trabajo sobre esta mutación el que determinó que las mutaciones son aleatorias.
Los ribosomas mutantes resistentes a los antibióticos tienen mayor información de Spetner
En términos de la métrica de unión de Spetner, la proteína mutante S12 es más específica; ahora solo se une al tRNA en lugar de al tRNA y la estreptomicina. Ahora, la estreptomicina no es un sustrato, por lo que podrías objetar el uso de la especificidad de unión en este caso. Sin embargo, la unión de estreptomicina es el propio ejemplo de Spetner para la especificidad de unión, y la utiliza como ejemplo de un sistema de unión "llave y cerradura" (véase arriba). Recuerda también que su métrica es generalmente aplicable a todas las interacciones ligando-receptor (recuerda que en su ejemplo de "enzima palabra", solo está ocurriendo unión, por lo que, según sus propios ejemplos, el ligando no tiene que ser un sustrato).
Además, la mutación sí tiene un efecto en la precisión de unión al sustrato del ribosoma. Aunque la estreptomicina se une en un sitio diferente al sitio real de corrección de pruebas del tRNA, este es un ejemplo clásico de algo llamado modulación alostérica. En el modelo simple de Spetner sobre la unión de sustratos, solo se consideran los ligandos que se unen directamente al sitio activo. Sin embargo, muchas enzimas y receptores hormonales tienen sitios para ligandos distintos al sitio activo, donde la unión de ligandos a estos sitios distantes resulta en la modificación de la unión de sustrato/ligando en el sitio activo. Estos tipos de ligandos se llaman moduladores alostéricos y son muy importantes en biología. Incluyen la unión de sodio a un bolsillo en la enzima digestora de proteínas tripsina, lejos del sitio activo, lo cual altera la actividad catalítica de la tripsina y la unión de glicina a un sitio modulador lejos del sitio de unión de glutamato en el receptor de glutamato (lo cual altera la actividad nerviosa) y muchos otros ejemplos. Si excluye de la consideración un modulador alostérico de la acción enzimática como la estreptomicina, entonces está ignorando la biología.
Los ribosomas que se unen a la estreptomicina producen proteínas defectuosas porque la estreptomicina altera el centro de corrección de pruebas (que es cómo la estreptomicina mata a las bacterias). La versión mutante que no se une a la estreptomicina es en realidad MÁS precisa, es decir, más ESPECÍFICA, que el tipo salvaje. El centro de corrección de pruebas del tipo salvaje comete algunos errores incluso en ausencia de estreptomicina, y las formas mutantes cometen aún menos errores que el tipo salvaje (aproximadamente 85% menos; 12).
Este es un aumento claro en la especificidad de unión de Spetner:
- El producto génico mutante no se une a la estreptomicina en absoluto (tiene un ligando en lugar de dos)
- Se une al sustrato peptidil tRNA con mayor precisión
- Cataliza la síntesis de péptidos más precisa.
¿Spetner reconoce esto? No, Spetner ahora cambia a la medida de expectativa y afirma (sin evidencia) que, dado que debe haber más secuencias S12 que no se unen a la estreptomicina que las que sí lo hacen, la información debe haber disminuido (¿por qué no realizó este análisis sobre la enzima ribulosa?).
Sin embargo, está completamente equivocado. El conjunto de todos los genes rsPL que producen la unión de estreptomicina a S12 es de alrededor de 1060. El conjunto de todos los genes rsPL que no unen estreptomicina es también de alrededor de 1060 (según un análisis de mutaciones neutras utilizando el método de Yockey (13)). Por lo tanto, la diferencia de información es tan pequeña que es inexistente. Si solo consideramos los cambios de aminoácidos en los aminoácidos de unión a estreptomicina, entonces hay aproximadamente el mismo número de sustituciones que permitirán la unión que aquellas que no la permitirán (10 resistentes frente a 6 normales, véase 14).
Importante, existe una mutación particular, AAA42 -> AGA42 (lisina a arginina) que no se une a la estreptomicina Y tiene precisión y tasas de traducción de tipo salvaje (15). Esta es la única mutación posible (simple) que hace esto, por lo que su conjunto es más pequeño que el conjunto de tipo salvaje. Otro ejemplo es la mutación que resulta en la subunidad S12 que aún se une a la estreptomicina y es resistente a la estreptomicina. De nuevo, esta es la única mutación posible, por lo que, según la medida de expectativa, esta mutación tiene más información que el tipo salvaje.
Por lo tanto, vemos que, sin importar cómo Spetner aplique sus métricas, la mutación resistente a la estreptomicina tiene más información que el gen de tipo salvaje.
Spetner y las mutaciones "dirigidas": [Inicio]
Spetner cree haber demostrado que las mutaciones aleatorias no pueden producir los aumentos en la información que serían necesarios en la evolución (como hemos visto, se equivoca). Spetner propone que la evolución ocurre mediante mutaciones "dirigidas" (con la implicación de que la Inteligencia Divina está de alguna manera detrás de esta "dirección"). La aparente existencia de mutaciones "dirigidas" se basó en algunos trabajos tempranos de Barry Hall. Sin embargo, Spetner no siguió la investigación más reciente sobre este trabajo y malinterpreta el origen y la significación de las mutaciones "dirigidas".
La mayoría de la gente sabe que las mutaciones surgen al azar en las células que se dividen, y que las mutaciones ocurren en diversos lugares del ciclo de replicación como consecuencia de daños (por ejemplo, de químicos mutagénicos o radiación) o de errores de corrección de pruebas en el proceso de copia.
Sin embargo, la mayoría de la gente no es consciente de que existe un cambio significativo de ADN en células no en crecimiento (no divididas) (16).
Las mutaciones adaptativas (dirigidas) son mutaciones que aparentemente dan lugar a la aparición selectiva de mutaciones favorables. La denominación de estas mutaciones ha causado considerable controversia y han sido llamadas mutaciones adaptativas, dirigidas, cairnsianas, inducidas por selección y mutaciones asociadas al estilo de vida estresante (SLAM). Un investigador acuñó el nombre "Fred" al intentar encontrar un nombre que no inflamara a los críticos, y "Fred" parece haber encontrado su camino al menos en el discurso informal de los investigadores relevantes (17).
"Fred" solo ocurre bajo selección no letal en células no divididas, y se ha sugerido que es un mecanismo neo-lamarckiano para introducir información ambiental en los genes. Sin embargo, "Fred" no es tal cosa (16,17,18), y no está dirigido en el sentido que sugiere Spetner.
- Fred no es lamarckiano. No está involucrada ninguna transcriptasa inversa; los mutantes no se transcriben de nuevo desde alguna proteína alterada por el medio ambiente ni siquiera desde un ARNm modificado por azar.
- Fred no está dirigido; las mutaciones se encuentran de forma aleatoria en todo el genoma, no solo en el gen "adaptativo".
- Fred depende de la recombinación; dañar cualquiera de las enzimas de recombinación RecA o RecBCD reduce la tasa de mutaciones "adaptativas".
- Fred depende de la ADN polimerasa; en mutantes de ADN pol III con mejor corrección de pruebas, la tasa de mutaciones "adaptativas" se reduce.
- Fred depende en gran medida de la reparación de desajustes defectuosa (MMR). Los defectos en MMR aumentan las mutaciones "adaptativas" y un MMR aumentado o la sobreexpresión de MMR reduce las mutaciones "adaptativas". Crucialmente, MMR se reduce en células no crecientes y estresadas.
Por lo tanto, el siguiente modelo muestra cómo ocurren las mutaciones "adaptativas". Las células no crecientes privadas de nutrientes están bajo estrés; el estrés provoca roturas de doble cadena en el ADN, la recombinación mediante RecBCD inicia la síntesis de ADN, la ADN PolIII completa el trabajo pero comete errores, los cuales pasan desapercibidos porque el hambre ha desactivado en gran medida la reparación de errores. Esto resulta en mutaciones a nivel del genoma a una tasa más rápida que la normal, y la ocasional mutante que puede utilizar el sustrato "selectivo" (16,17,18).
El ejemplo canónico es E. coli que tienen un gen Lac dañado y no pueden utilizar lactosa; cuando se siembran en un medio que tiene solo lactosa como fuente de carbono, las células no pueden crecer, pero después de un tiempo aparecen colonias que pueden utilizar lactosa. Estas colonias contienen una versión del gen dañado que ha sido restaurada a la función (generalmente mediante un desplazamiento de marco de 1 pb (16,17,18)).
Así que Fred, aunque ciertamente emocionante desde el punto de vista genético, resulta ser solo otra mutación aleatoria aburrida.
Conclusión: [Inicio]
En resumen, aunque los argumentos de Spetner son superficialmente plausibles, un análisis más profundo con algún conocimiento de bioquímica revela fallos masivos. Spetner se equivoca en los detalles de la biología: la especificidad del ligando no está directamente gobernada por la longitud de la cadena de unión como exige la teoría de Spetner, y la unión del ligando no es un "asunto de todo o nada". Esto invalida sus análisis. Incluso entonces, los propios ejemplos de Spetner no respaldan sus afirmaciones. Además, al utilizar sus métricas, Spetner cambia las métricas cuando se muestran cambios inconvenientes.
Referencias: [Inicio]
1. Spetner, L. M. (1998) ¡NO POR SUERTE! Desmintiendo la teoría moderna de la evolución, Judaica Press, Nueva York.
2. Shannon CE (1948) Una Teoría Matemática de la Comunicación. Bell Sys Techl J 27.
3. Schneider TD, (2000) Evolución de la información biológica, Nucleic Acids Res, 28(14): 2794-2799.
4. Max E (2001) La evolución de la aptitud mejorada
5. Hunyady et al., (2003) Inducción de agonistas y selección conformacional durante la activación de un receptor acoplado a proteínas G. [ PubMed] TIPS 24, 81-86
6. Le MT et al., (2002) La angiotensina IV es un potente agonista para los receptores AT1 humanos constitutivamente activos. J Biol Chem, 277, 23107-23110
7. Knowle D, et al., (2001) Papel del Asp297 del receptor AT2 en el enlace de alta afinidad a diferentes ligandos peptídicos. [ PubMed] Peptides, 22, 2145-2149
8. Turner, CA, et al., (1999) Papel del His273 ubicado en el sexto dominio transmembrana del subtipo de receptor de angiotensina II AT2 en la interacción ligando-receptor. [ PubMed] BBRC, 257, 704-707
9. Hartley, B.S. (1984) Evolución experimental de la ribitol deshidrogenasa. En R.P. Mortlock (ed.), "Microorganismos como sistemas modelo para estudiar la evolución" (pp.23 - 54) Plenum, Nueva York.
10. Matsumura I, Ellington AD. La evolución in vitro de la beta-glucuronidasa hacia una beta-galactosidasa procede a través de intermediarios inespecíficos. [ PubMed] J Mol Biol. 2001 Jan 12;305(2):331-9
11. Hayashi Y, et al., ¿Puede una secuencia arbitraria evolucionar hacia la adquisición de una función biológica? [ PubMed] J Mol Evol. 2003 Feb;56(2):162-8.
12. Björkman J, et al., Nuevas mutaciones ribosomales que afectan la precisión de la traducción, la resistencia a los antibióticos y la virulencia de Salmonella typhimurium. Mol Microbiol. 1999 Jan;31(1):53-8.
13. Yockey HP. (1992) Teoría de la información y biología molecular, Cambridge University Press. Capítulo 6.3
14. Tolvonen JM et al., Mol. Micro. 1999, 1735-1746
15. Björkman J et al., (1998) Virulencia de Salmonella typhimurium resistente a los antibióticos. Proc Natl Acad Sci USA. 95, 3949-3953.
16. Foster PL & Rosche WA. Mecanismos de mutación en células no divididas. Perspectivas del estudio de la mutación adaptativa en Escherichia coli [ PubMed] Ann N Y Acad Sci 1999, 870, 133-45.
17. Rosenberg, SM. Mutación para la supervivencia [ PubMed] Curr Opinion Gen Dev 1997, 7:829-834
18. Rosche WA, y Foster PL. (1999 Jun 8). El papel de los hipermutadores transitorios en la mutación adaptativa en Escherichia coli. Proc Natl Acad Sci U S A , 96, 6862-7.
Agradecimientos:
Muchas gracias a Chris Ho-Stuart, Michael Hopkins, Bill Hudson y Douglas Theobald por sus sugerencias útiles y la corrección de pruebas.