Otros enlaces:
|
La historia de los embriones de Haeckel es diferente de una manera importante a la de los otros capítulos en el libro de Jonathan Wells. Como muestran los otros autores, Wells ha distorsionado ideas que son fundamentalmente verdaderas para hacer su punto: toda su retórica en contrario, Archaeopteryx es un fósil transicional, las polillas peppered y los pinzones de Darwin dicen cosas significativas sobre la evolución, las moscas de cuatro alas dicen cosas significativas sobre las vías de desarrollo, y así sucesivamente. En esas partes del libro, Wells tiene que tratar de ocultar una verdad mediante la mala interpretación y la tergiversación de ella.
En el caso de Haeckel, sin embargo, debo comenzar admitiendo que Wells tiene razón en lo esencial de la historia. Haeckel se equivocó. Su teoría era inválida, algunos de sus dibujos fueron falsificados e interpretó deliberadamente los datos de manera excesiva para respaldar una tesis falsa. Además, fue influyente tanto en las ciencias como en la prensa popular; su teoría sigue siendo citada en esta última hoy en día. Wells también tiene razón al criticar a los autores de los libros de texto por perpetuar el infame diagrama de Haeckel sin comentar sus inexactitudes o la forma en que fue mal utilizado para apoyar una teoría falsificada.
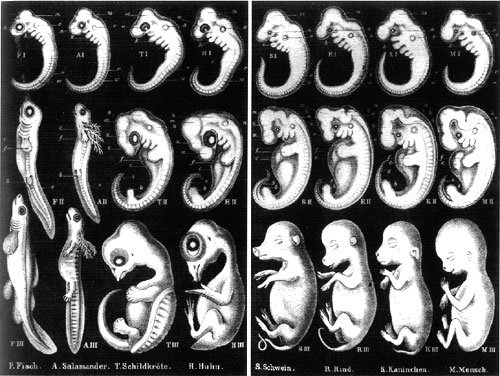
Desafortunadamente, lo que Wells intenta hacer en este capítulo es tomar esta teoría inválida y desacreditada y manchar con ella la biología evolutiva moderna (e incluso no tan moderna). La ley biogenética no es darwinismo ni neodarwinismo, sin embargo. No forma parte de ninguna teoría evolutiva moderna. Wells está llevando a cabo un engaño aquí, reuniendo la evidencia y las citas que debidamente demuestran la falsedad del dogma haeckeliano, y luego afirmando que esto forma parte de "nuestra mejor evidencia para la teoría de Darwin".
¿Quién fue Haeckel y cuál fue su teoría?
Ernst Haeckel (1834-1919) fue un científico alemán extremadamente influyente. Tenía una reputación envidiable como embriólogo comparativo, pero su principal reclamo a la fama fue que fue un adoptante temprano de la teoría de Darwin que utilizó la evidencia de la embriología para apoyar la evolución. Wells tiene toda la razón al mencionar la importancia de Haeckel en la biología del siglo XIX. No hay duda de que sus esfuerzos por popularizar la teoría fueron importantes para dar credibilidad a la evolución en el establecimiento científico, y también para los profanos. Darwin y Haeckel se conocieron y correspondieron, y cada uno influyó fuertemente en las teorías del otro. Sin embargo, las teorías de Haeckel debían una deuda aún mayor a una tradición filosófica anterior, en particular la obra de Goethe y Lamarck. Aunque Darwin fue apreciativo del apoyo de Haeckel a la selección natural, también fue cauteloso al utilizar las ideas de Haeckel en sus escritos; Darwin se basó mucho más en los datos embriológicos de von Baer para apoyar la descendencia común.
Además, la teoría de Haeckel estaba podrida en lo más profundo. Era incorrecta tanto en principio como en el conjunto de observaciones sesgadas y manipuladas utilizadas para sostenerla. Esto fue una tragedia para la ciencia, porque llevó a los biólogos evolutivos y a los biólogos del desarrollo a un callejón sin salida, provocando una desafortunada separación entre los campos del desarrollo y la evolución que solo recientemente ha sido corregida.
La teoría de Haeckel se resume en su memorable aforismo, "la ontogenía recapitula la filogenia", también llamado la ley biogenética. Lo que esto significa es que el desarrollo (ontogenía) repite la historia evolutiva (filogenia) del organismo: si evolucionamos de un pez que evolucionó en un reptil que evolucionó en nosotros, nuestros embriones reflejan físicamente esa historia, pasando por una etapa similar a la de un pez y luego a una etapa similar a la de un reptil. ¿Cómo podría ocurrir esto? Él argumentó que la historia evolutiva era literalmente la fuerza impulsora detrás del desarrollo, y que las experiencias de nuestros ancestros estaban físicamente escritas en nuestro material hereditario. Esto fue una extensión lógica de su creencia en la herencia lamarckiana, o la herencia de caracteres adquiridos. Si la actividad de un organismo puede imprimirse en su genética, entonces el desarrollo podría ser simplemente un resumen de las actividades de los padres, abuelos y antepasados cada vez más remotos. Esta fue una idea extremadamente atractiva para los científicos; es como si el desarrollo fuera una máquina del tiempo que les permitiera mirar hacia el pasado lejano, simplemente estudiando las etapas tempranas del desarrollo.
Desafortunadamente, también estaba completamente equivocado.
Los descubrimientos que finalmente demolieron los premisas subyacentes de la ley biogénica fueron los principios de la genética y las observaciones empíricas de embriones. La herencia lamarckiana simplemente no ocurre, como todos aprendimos en biología de secundaria. El ADN es el agente de la herencia, y no se modifica por nuestras acciones ordinarias - si usted se hace un tatuaje, no se escribe también en los cromosomas de sus espermatozoides o óvulos, y no hay riesgo de que sus hijos nazcan con "Mom" grabado en su brazo. El descubrimiento de que Haeckel había tomado atajos inexcusables con sus ilustraciones fue un problema relativamente menor para su teoría, porque la tendencia general de sus observaciones (que los embriones de vertebrados se parecen fuertemente entre sí) había sido confirmada independientemente. Lo que realmente hundió toda la teoría fue que su fundamento fue eliminado, lo que hace que la afirmación de Wells de que los biólogos están intentando "resucitar la recapitulación" sea bastante peculiar, ya que no hay muchos que vean alguna validez en la herencia lamarckiana.
Wells también querría que el lector pensara que los embriones de Haeckel eran una pieza clave del rompecabezas que Darwin mismo utilizó para ensamblar su teoría, y que por lo tanto el darwinismo se construyó sobre una base falsa. Sin embargo, omite mencionar un asunto importante de cronología. El Origen de las Especies de Darwin fue publicado en 1859, después de décadas de investigación. La teoría de Haeckel fue publicada por primera vez en 1866, y los diagramas de Haeckel fueron publicados en 1874. Haeckel no tuvo ninguna aportación significativa al Origen, pero Darwin (y Lamarck y un número de filósofos alemanes, como Goethe) influyeron profundamente en Haeckel. Claramente, las posteriores elaboraciones de Darwin sobre la teoría de la evolución fueron complementadas por las ideas de Haeckel, en particular sus ideas sobre la herencia, pero son precisamente esos componentes del darwinismo que fueron invalidados por la genética mendeliana y la síntesis neodarwiniana.
¿Qué es la etapa filotípica?
Darwin dijo: "Pocos puntos me dieron tanta satisfacción cuando trabajaba en El Origen como la explicación de la amplia diferencia entre el embrión y el animal adulto en muchas clases, y la estrecha semejanza de los embriones dentro de la misma clase". A principios del siglo XIX, von Baer observó que los embriones de diferentes especies no podían distinguirse tan fácilmente entre sí como los adultos. Esta es una observación simple que ha sido realizada numerosas veces en los últimos cientos de años, una observación que no está ligada a ninguna teoría en particular, ya sea de creación o de evolución: von Baer realizó estas observaciones 30 años antes de que Darwin publicara, y no aceptó la evolución entonces ni después. Es decir, mientras que los adultos vertebrados pueden parecer muy diferentes entre sí, los embriones vertebrados pasan por un periodo en el desarrollo en el que todos se parecen más fuertemente entre sí. Este periodo se llama etapa filotípica. En este momento del desarrollo, los embriones vertebrados expresan un conjunto de caracteres comunes a toda la línea vertebrada: tienen un notocordio y un cordón nervioso dorsal, tienen arcos faríngeos y una cola, y tienen una serie repetitiva de bloques musculares llamados somitas. La mayoría de las características que distinguen a los diferentes grupos de vertebrados, como patas o aletas, pelo o plumas o escamas, y alas o antebrazos, aún no se han desarrollado. Además, ahora sabemos que este es el periodo durante el cual se expresa por primera vez un conjunto de genes cruciales para la formación de patrones (los genes emx/otx/Hox) y se establece el plano molecular del cuerpo.
La observación no está vinculada a ninguna teoría en particular, pero aún debemos explicar la existencia de este fenómeno en cualquier teoría del origen. Haeckel lo intentó. Su explicación fue un modelo defectuoso de la herencia que afirmaba que existían restricciones sobre las propiedades de la genética, de tal manera que solo las fases finales eran fácilmente modificables. Esto significaba que las nuevas características se añadían generalmente solo al final del programa de desarrollo, de modo que la secuencia de desarrollo era aproximadamente la misma que la secuencia de evolución. Sin embargo, esta explicación ha sido refutada. No solo la genética no funciona de esa manera, sino que el desarrollo no progresa de una forma que se parezca a la secuencia evolutiva; como señala Wells (repetiendo las mismas críticas que se han hecho durante más de un siglo), las etapas más tempranas del desarrollo pueden diferir radicalmente. Luego convergen en una apariencia similar en la etapa filotípica, y divergen nuevamente en morfología.
Las teorías modernas del desarrollo y la evolución proponen algo que se ajusta a las observaciones y que Wells no puede descartar fácilmente. Los genes pueden modificarse para actuar en prácticamente cualquier punto del desarrollo, por lo que la restricción teórica impuesta por Haeckel no existe. Las variaciones entre especies en las etapas más tempranas fueron un problema para Haeckel, pero no son en absoluto incompatibles con la biología del desarrollo moderna. Incluso no hay un requisito de identidad morfológica absoluta en la etapa filotípica. Como señala Wells, Michael Richardson ha estado identificando variaciones dentro de esa etapa entre especies.
En Íconos de la Evolución, Wells está golpeando un caballo muerto al atacar exclusivamente la explicación de Haeckel e ignorando las ideas modernas. Además, no está contribuyendo a nuestra comprensión de por qué los vertebrados se desarrollan de la manera en que lo hacen. ¿Por qué los animales que difieren en apariencia como adultos exhiben similitudes como embriones? ¿Por qué los embriones humanos tienen colas y construyen sus caras desde la misma base embrional que los peces utilizan para construir branquias? Estas son preguntas que Wells falla en abordar, y su enfoque parece ser intentar argumentar en su lugar que no exhiben similitudes como embriones. Sin embargo, sí lo hacen. Estas similitudes argumentan a favor de la descendencia común.
¿Qué hay de las diferencias tempranas en el desarrollo?
Wells hace mucho énfasis en el hecho de que la ley biogenética presupone que los embriones deberían ser más similares en la etapa de desarrollo más temprana posible, sin embargo, el período de mayor similitud, la etapa filotípica, ocurre mucho más avanzado en el desarrollo, y que las etapas más tempranas exhiben diferencias aparentemente significativas. Esto es cierto, pero irrelevante.
- La ley biogénica no es darwinismo. Cuestionar la ley biogénica es fácil de hacer; se ha hecho repetidamente en la literatura científica y no daña en absoluto a la biología evolutiva moderna.
- Esta observación no es noticia. Von Baer señaló en la década de 1820 que los tejidos más tempranos en diferenciarse en el embrión mamífero son las membranas extraembrionarias, las cuales también resultan ser entre las características más recientes en la línea evolutiva de los mamíferos. La figura que Wells usa para hacer este punto (Figura 5-3) es una versión redibujada de una figura similar publicada por Elinson en 1987.
- Wells minimiza las similitudes que existen para enfatizar las diferencias. Este es el mismo juego sesgado que jugó Haeckel: la única diferencia es que él enfatizó las similitudes en lugar de las diferencias.
Es necesario explicar con más detalle ese último punto. Lo que la biología del desarrollo moderna ha ido descubriendo es que estas etapas iniciales en los embriones de diferentes vertebrados son sustancialmente similares. Las diferencias son superficiales y, en muchos casos, una consecuencia de diferentes cantidades de inversión materna. Por ejemplo, una de las primeras tareas de un huevo fecundado y unicelular es dividirse repetidamente para producir un gran número de células. En animales que tienen una gran cantidad de yema, como un pez, un reptil o un ave, la yema es demasiado grande para ser dividida eficientemente, por lo que la célula se divide parcialmente, produciendo una capa de células sobre la yema. En animales que tienen relativamente poca yema bien dispersa, como la rana, o que no tienen yema en absoluto, como en los mamíferos, el huevo puede dividirse completamente y producir una bola de células. Esa es la magnitud de la diferencia, ya sea que tengas una masa de células por sí misma o una masa extendida sobre una yema.
Estas diferencias en la forma conducen a algunas diferencias aparentes en la siguiente etapa del desarrollo, la gastrulación. La función crítica de la gastrulación es que algunas células deben moverse hacia el interior, dentro de la masa de células. Este movimiento de láminas de células genera la estructura tridimensional del embrión y, durante el proceso, ofrece a las células oportunidades para interactuar e inducir nuevos tipos de tejido. Sin embargo, ya sea que el embrión sea una bola de células o una masa sobre un vitelo, todos los vertebrados realizan movimientos equivalentes durante la gastrulación; nuevamente, las diferencias son superficiales, dependiendo de si el grupo de células está en forma de bola o aplanado. Existen otras diferencias que pueden catalogarse, como si las células se mueven hacia el interior como una lámina coherente y adhesiva o si se delaminan y se mueven hacia el interior como células individuales, pero nuevamente, el proceso central, que es el movimiento hacia el interior para establecer nuevas capas, es el mismo.
A un nivel más fundamental, parece que los vertebrados comparten un conjunto común de moléculas de señalización, como BMP-4 y chordin, que son importantes para definir la polaridad y especificar tipos de tejido en estas etapas tempranas. Los procesos de señalización celular importantes para inducir nuevos tejidos durante la gastrulación son similares en todos los vertebrados, y las mismas respuestas están surgiendo en peces y ratones, a pesar de las diferencias morfológicas en sus estructuras. Es engañoso centrarse en las aparentes diferencias de forma a un nivel general, cuando los aspectos más profundos, moleculares, de estas etapas del desarrollo son homólogos.
¿Qué hay de malo con las "hendiduras branquiales"?
Wells está bastante descontento con el término común "hendiduras branquiales". Dedica varias páginas diciéndonos que los mamíferos embrionarios no tienen branquias, y que incluso en las etapas en las que los peces tienen hendiduras branquiales, no tienen branquias. Sin embargo, está perdiendo el punto: nadie ha afirmado que tengan branquias. Tienen "hendiduras branquiales", algo completamente diferente. Se ha dejado envolver tanto en un argumento etimológico trivial que ha perdido toda noción de la realidad biológica.
Los embriones de vertebrados universalmente poseen estructuras prominentes en su región cervical que son llamadas por diversos nombres en la literatura científica: branquiales, faríngeos, o bolsas, surcos o hendiduras viscerales o arcos. Debido a que pueden aparecer como una serie repetida de hendiduras en el cuello del embrión, parecida al patrón de elementos repetidos en el cuello de los peces adultos, también han sido coloquialmente llamadas "hendiduras branquiales" o "bolsas branquiales". No son, sin embargo, branquias - y los científicos no han estado afirmando que lo sean (Wells incluso cita a varios autores, Wolpert y Rager, quienes explicitan este hecho simple y obvio). Entonces, ¿qué son?
Las "hendiduras branquiales" son elementos estructurales comunes del desarrollo craneofacial de los vertebrados. "Común" es el término importante aquí. Resulta que todos los vertebrados construyen su rostro de la misma manera, algo algo improbable y contraintuitivo; es esta profunda similitud la raíz del argumento evolutivo de que refleja la descendencia común.
La cabeza de todos los embriones de vertebrados, ya sean peces o humanos, puede describirse simplemente como un tubo curvado compuesto principalmente por tejido presumiblemente cerebral (Figura 2), con una serie de 4 a 7 tejidos en forma de dedos que cuelgan de él, los arcos faríngeos. Lo que consideramos rostro, todo desde justo debajo de los ojos, hasta las orejas y hacia abajo hasta el cuello, está ausente. En su lugar, tenemos estos bultos colgantes, cada uno de los cuales contendrá una varilla cartilaginosa, una columna muscular, una rama significativa del sistema circulatorio y una variedad de otros tipos celulares. Estos arcos son módulos reiterados que posteriormente se fusionarán y reorganizarán (junto con otros tejidos craneales, y más importante aún, una población migratoria de células desde la parte superior de la cabeza llamada cresta neural) para formar el rostro más familiar. Lo hacen de manera similar en todos los vertebrados: el primer arco faríngeo, por ejemplo, siempre forma la mandíbula, y el segundo arco siempre forma el hioides. También existen diferencias que surgen en diferentes clases. Partes de los dos primeros arcos encuentran su lugar en los huesos del oído mamífero. Los terceros y arcos subsiguientes en los peces terminan en las branquias, mientras que esos mismos arcos en un humano forman una serie de cartílagos en la garganta. El tercero se fusiona con el hioides, el cuarto forma una parte importante del cartílago tiroides, y el quinto forma los cartílagos cricoides y aritenoides. Los elementos no cartilaginosos de estas estructuras terminan incorporados en todo tipo de tejidos, glándulas y músculos y epitelios, del cuello y el rostro.
 |
Este patrón común de desarrollo es algo genuinamente notable. Es una de las razones por las que los primeros biólogos evolutivos, como Charles Darwin, argumentaron que la evidencia de la embriología era tan importante para su teoría: vemos que tales procesos intrincados se repiten una y otra vez de maneras tan similares en especie tras especie. Dado que los arcos faríngeos iniciales se reorganizan radicalmente a lo largo del desarrollo, no hay una razón obvia por la que todos los embriones de vertebrados comiencen con estructuras virtualmente idénticas que están igualmente distantes de su morfología final, más allá de que reflejen una base morfológica compartida y un ancestro común.
Wells quiere negar todo esto. Para ello, se centra exclusivamente en la terminología. Señala que las "hendiduras branquiales" no son branquias, un hecho bien comprendido por cada biólogo del desarrollo. Dice que "nunca son 'parecidas a branquias' excepto en el sentido superficial de que forman una serie de líneas paralelas en la región del cuello". Esto es cierto, y nadie atribuye ninguna significación más profunda al término que esa. Sin embargo, Wells luego intenta afirmar que "la única forma de ver estructuras 'parecidas a branquias' en embriones humanos es leer la evolución en el desarrollo" y que "no hay forma de que las 'hendiduras branquiales' en embriones humanos puedan lógicamente servir como evidencia para la evolución". Estas afirmaciones son falsas y engañosas. Como él mismo ha señalado, una forma de ver estructuras "parecidas a branquias" en embriones es observar que forman un patrón repetido de líneas en el cuello; eso no presupone la evolución y es una fuente legítima para el nombre. Por supuesto, entonces tenemos que preguntarnos por qué los embriones humanos tienen estas extrañas líneas, y por qué todos los demás vertebrados también las tienen... y esta comúnidad de estructura es la evidencia para la evolución, no las implicaciones superficiales de los nombres.
Todos los embriones de vertebrados tienen arcos faríngeos. El hecho de que a veces estas estructuras se refieran con el término más simple "hendiduras branquiales" en los libros de texto no significa que no existan o que desaparezca la homología, aunque ese es el juego que Wells querría jugar.
¿Dónde están todos los recapitulacionistas?
Una premisa que Wells plantea con regularidad en este capítulo es que los biólogos evolutivos se han basado en las falsas doctrinas de Haeckel para sostener el dogma darwinista. Afirma que las teorías haeckelianas "han surgido periódicamente, como un fénix, de las cenizas de la refutación empírica" a lo largo del siglo XX. Utiliza títulos de sección como "Resucitando la recapitulación" y "Haeckel está muerto. ¡Viva Haeckel!". Acusa a los biólogos de una conspiración del silencio, ocultando los defectos del trabajo de Haeckel por un lado, y utilizándolo como evidencia para la evolución por el otro.
Nada de esto es cierto.
Esto es tan patentemente obvio que solo necesitamos utilizar el propio trabajo de Wells para demostrarlo. Él cita a varios autores que discuten a Haeckel o la ley biogenética:
- Adam Sedgwick, 1894
- William Garstang, 1922
- Gavin de Beer, 1958
- William Ballard, 1976
- Stephen J. Gould, 1977
- Richard Elinson, 1987
- Jane Oppenheimer, 1987
- Michael Richardson, 1995
- Stephen J. Gould, 2000
Sin embargo, aquí está lo sorprendente: todos estos autores condenan la idea de que el desarrollo embrionario sigue el patrón evolutivo en términos inequívocos! Sedgwick, por ejemplo, compiló una extensa lista de objeciones a la recapitulación tal como fue formulada por von Baer y Haeckel, y la rechazó específicamente como insostenible - en 1894. Esto representa más de un siglo de denegación inequívoca de la recapitulación. La fecha puede retroceder aún más, ya que von Baer publicó su crítica de las interpretaciones recapitulativas de sus observaciones en 1828. Wells cita a estos autores para respaldar su argumento de que las ideas de Haeckel son falsas, pero al mismo tiempo intenta fingir que los biólogos evolutivos son todos fans ocultos de Haeckel. Si ese es el caso, ¿dónde están las citas a científicos modernos prominentes que defienden sus teorías?
El caso de Stephen J. Gould ilustra particularmente bien la estrategia de doble cara de Wells. En 1977, Gould escribió un excelente libro académico sobre la historia entrelazada de la embriología y la evolución, titulado Ontogeny and Phylogeny. Como se podría deducir del título, Haeckel es un personaje prominente en el libro, y sus teorías y sus consecuencias en el campo son disecadas en detalle y sin piedad. Gould también volvió a este tema en su columna en la revista Natural History en 2000, centrando su comentario en el descubrimiento de una crítica mordaz de Haeckel por uno de sus contemporáneos, Louis Agassiz. Gould también ha escrito otros artículos relacionados, despreciando a los autores de libros de texto por su lamentable hábito de reciclar texto y figuras mucho más allá de lo razonable. Sin embargo, ¿cómo distorsiona Wells los hechos? Él afirma que Gould "...(a pesar de haber conocido la verdad durante más de veinte años) mantuvo la boca cerrada hasta que un "creacionista" (en realidad, un colega biólogo) [se refiere aquí a Michael Behe] expuso el problema"!
Este es un ejemplo notable del tipo de doble discurso deshonesto utilizado por Wells a lo largo de su libro. Puede citar autor tras autor que claramente condenan las ideas de Haeckel entre 1894 y 2000, y al mismo tiempo afirmar que los biólogos han estado "resucitando el recapitulacionismo" a lo largo del siglo XX. Afirma que Behe "expuso el problema", pero Behe no ha realizado ninguna investigación en embriología, no ha publicado nada original sobre la historia de la ciencia del siglo XIX y no merece crédito aquí: este "problema" fue expuesto hace más de un siglo, y el verdadero trabajo creativo e innovador en el análisis embriológico que se realiza aquí es de un evolucionista, Michael Richardson, no de Behe. También me sorprende que Gould pueda publicar un libro de casi 500 páginas que sea bien recibido críticamente y frecuentemente citado, y aún así ser acusado de mantenerse callado sobre el tema.
Gould no está solo. El autor de uno de los libros de texto más influyentes en biología del desarrollo, Scott Gilbert, describió la ley biogenética como "desastrosa" e "insostenible". Raff y Kaufman hicieron el mismo caso contra Haeckel que Wells hace aquí, por lo que lo que Wells nos enseña no sorprende a nadie.
¿Son realmente tan malos los libros de texto?
Wells está particularmente enfurecido con los autores de libros de texto introductorios que, según él, están engañando a sus estudiantes. Estoy de acuerdo en que puede argumentar razonablemente que los libros de texto no deberían utilizar los dibujos obsoletos e inexactos realizados después del trabajo de Haeckel, pero, en lo que considero la línea más divertida de este capítulo entero, Wells expresa indignación de que "Algunos libros de texto, en lugar de reproducir o redibujar los embriones de Haeckel, utilizan fotos reales". ¡Cómo se atreven aquellos nefastos autores de libros de texto a utilizar datos fotográficos para apoyar sus ideas!
En su primer apéndice, Wells califica una serie de libros de texto universitarios, asignándoles calificaciones de letras según qué tan bien sus ideas se ajustan a las suyas. Todos los libros reciben una D o una F por su tratamiento de los embriones de vertebrados. Sin embargo, es evidente que está manipulando las reglas en su contra.
Para obtener una calificación de F, un libro de texto solo necesita utilizar una versión de los dibujos de Haeckel, afirmar que las similitudes tempranas en los embriones de vertebrados son evidencia de descendencia común y hacer referencia a las hendiduras branquiales. Estoy de acuerdo en que los libros no deberían utilizar las ilustraciones de Haeckel, a menos que sea para hacer un punto histórico. Sin embargo, las similitudes en la estructura son evidencia de descendencia común; es un poco injusto penalizar a un libro por hacer un punto válido. Del mismo modo, los embriones de vertebrados tienen "hendiduras branquiales", incluso si las asociaciones de ese término confunden al Dr. Wells.
Para mejorar su calificación a un D, un libro utiliza "fotografías engañosas" de embriones reales en lugar de los dibujos haeckelianos. Sin embargo, ¿cuál es la objeción aquí? Estos no son dibujos manipulados o inexactos. Son fotografías de embriones de vertebrados que ilustran con precisión cómo se ven, algo que cualquiera con un microscopio y acceso a embriones puede observar (Figura 3). ¿Será el siguiente en su agenda condenar las clases que permiten a los estudiantes examinar embriones vivos o preservados en el laboratorio?
 |
¿Qué tiene que hacer un libro de texto para obtener una A? Primero, no debe utilizar ilustraciones o fotografías engañosas. Wells no ha demostrado que las fotografías utilizadas en ninguno de estos libros sean engañosas en absoluto. Segundo, no debe utilizar el término "hendiduras branquiales". Este es un requisito arbitrario; no hay nada incorrecto en sí mismo con ese término en particular.
Tercero, debería discutir las diferencias en las etapas más tempranas (lo cual es una buena idea, pero puede ser impráctico en un texto introductorio) y llamar a esto un problema para la evolución darwiniana. Sin embargo, existe una dificultad para cumplir con este último requisito. Las diferencias pueden ser un problema para un modelo evolutivo haeckeliano, pero no lo son para las formulaciones modernas de la evolución. Un libro que sugiriera que las diferencias tempranas son un problema estaría forzando una interpretación errónea de la evidencia.
Finalmente, su cuarto requisito es que el libro deba considerar errónea la teoría de Darwin sobre los orígenes de los vertebrados. Presumo con esto que debe utilizar observaciones de embriones de vertebrados para argumentar de alguna manera que los diferentes vertebrados no evolucionaron de un ancestro común. Desafortunadamente, esta conclusión no se deriva de la evidencia. Las profundas similitudes en el proceso de desarrollo de todos los vertebrados apoyan la idea de que forman parte de una linaje común; además, los avances en la biología molecular del desarrollo han confirmado constantemente homologías aún mayores; simplemente no es una opción siquiera sugerir que estos organismos no tuvieron un ancestro común.
Uno de los libros de texto por los que Wells otorga una calificación de "D" es uno del que he impartido clases durante varios años, el Biology de Campbell. Revisé detenidamente la sección relevante de este libro en particular. Inmediatamente encontré una seria deficiencia: esta sección tiene solo dos párrafos de longitud, con una figura. Sería bueno ver más sustancia sobre este tema, especialmente en un libro que tiene 1175 páginas, pero un problema recurrente en los libros que Wells ha dirigido es que estos son repaso de toda la biología, desde la bioquímica hasta la ecología, por lo que no es demasiado sorprendente que todo reciba menos profundidad de tratamiento de la que nos gustaría. Sin embargo, no encontré nada en lo que Campbell ha escrito que sea objetable. Comienza con el punto de que "los organismos estrechamente relacionados pasan por etapas similares en su desarrollo embrionario", e ilustra esto con una fotografía de un embrión aviar y mamífero. Esta afirmación es correcta, y la figura respalda el punto. Termina la sección corrigiendo explícitamente las ideas de Haeckel, diciendo que "La teoría de la recapitulación es una exageración. Aunque los vertebrados comparten muchas características del desarrollo embrionario, no es como si un mamífero primero pasara por una 'etapa de pez', luego una 'etapa de anfibio', y así sucesivamente. La ontogenia puede proporcionar pistas sobre la filogenia, pero es importante recordar que todas las etapas del desarrollo pueden modificarse a lo largo de la evolución." Esto es totalmente correcto. No veo ningún errores de hecho en el tratamiento del tema por parte de Campbell, aunque sí creo que es lamentable que tan poco espacio pueda ser reservado para ello.
Esto no sugiere que los libros de texto estén exentos de defectos. Estoy de acuerdo con Wells en que las contorsiones de Balinski en su texto de 1975 para racionalizar la ley biogenética son lamentables. También concuerdo en que los libros de texto que ya están dando poca importancia a este tema particular no deberían estar desperdiciando espacio con la ilustración de Haeckel, que no es buena información biológica y solo tiene interés histórico. Wells va demasiado lejos, sin embargo. Intenta usar una antigua (aunque influyente) distorsión para descartar una serie de observaciones perfectamente válidas en sus intentos de desacreditar algunas conclusiones lógicas que entran en conflicto con sus prejuicios.
¿Qué lección deberíamos aprender de los embriones?
Wells está enviando el mensaje equivocado en su discusión sobre los embriones de Haeckel. Quiere que sus lectores piensen que la biología ha estado encontrando crecientes discrepancias en los principios del desarrollo, discrepancias que desafían la evolución al echar duda sobre la unidad de la línea vertebrada. Nada podría estar más lejos de la verdad. La biología molecular, en particular, nos ha estado mostrando que hay un mayor grado de similitud entre especies del que se sospechaba anteriormente.
Los genes Hox, por ejemplo, ilustran esta relación. Se trata de un conjunto de genes reconocibles por varias características: todos comparten una secuencia común de 180 pares de bases llamada homeobox (de la cual deriva su nombre), son todas proteínas de unión al ADN que actúan para encender o apagar otros genes, y tienen una organización única. Todos están dispuestos en un orden en el cromosoma que coincide con el orden de su expresión: es decir, un gen Hox que se activa en la cabeza está en un extremo del grupo de genes, un gen Hox que se activa en la cola está en el otro extremo, y todos los genes intermedios están en un orden espacial correspondiente (Figura 4). Estos genes están involucrados en la formación del patrón del organismo, estableciendo el plan corporal general. Funcionan para especificar dónde debe formarse la cabeza, por ejemplo, o el cuello o la región lumbar. Curiosamente, estos genes se activan primero en la etapa filotípica. Una buena razón para la conservación de la morfología en la etapa filotípica es que es cuando estos genes deben interactuar entre sí y con otros genes para bosquejar el plano para el desarrollo posterior. Existen interacciones globales entre genes, y los genes pueden reutilizarse en varios lugares. Por ejemplo, se ha encontrado que varios tejidos inducen tejido lenticular, incluyendo el mesodermo cardíaco, y cambiar el momento de la diferenciación cardíaca podría afectar así la formación del ojo. La morfología específica del filotipo realmente representa una base literal sobre la cual procede el resto del desarrollo, y es resistente al cambio evolutivo, porque hay demasiados eventos posteriores que dependen de él.
 |
Todos los embriones de vertebrados utilizan estos genes Hox para definir su plan corporal. Un resultado sorprendente de los enfoques moleculares en el desarrollo es que parece que todos los animales multicelulares, desde gusanos hasta insectos, mamíferos y medusas, utilizan este mismo conjunto organizado espacialmente de genes que contienen homeobox para definir su estructura. Existen diferencias: los vertebrados parecen haber duplicado el conjunto varias veces, de modo que tienen cuatro copias del conjunto de genes Hox, mientras que Drosophila (en el que se identificaron por primera vez estos genes) tiene un solo clúster; los ratones y los humanos tienen 13 genes Hox en cada clúster, mientras que Drosophila tiene 9 y Hydra tiene 2. La universalidad de estos genes en el desarrollo animal ha llevado al concepto de zootipo, o forma que vincula a todos los animales, en lugar de solo los embriones de un solo filo. Aunque Jonathan Wells querría argumentar que existe una falta de correspondencia entre embriones dentro de solo la línea de los vertebrados, los biólogos moleculares han estado descubriendo información que amplía la correspondencia, encontrando similitudes fundamentales que vinculan a todos los animales.
Conclusión
Jonathan Wells desea desacreditar la evolución, y en los embriones de Haeckel, ha encontrado una historia a su gusto. Hay un poco de falsificación intencional en ello, hay una clara afiliación con Darwin mismo, y hay una larga historia de reconocimiento de la influencia de Haeckel entremezclada con una repudiación inequívoca de sus ideas. Todo lo que tiene que hacer es intentar entrelazar las teorías desacreditadas y la mala reputación moderna de Haeckel con el conjunto de observaciones válidas y las explicaciones modernas, y puede enterrar la verdad bajo insinuaciones y asociaciones. Sin embargo, solo tenemos que recordar tres cosas:
- La teoría de la evolución no se basa en las observaciones o teorías de Haeckel. El trabajo de Haeckel fue desacreditado en el siglo XIX y no ha sido relevante para la biología desde el redescubrimiento de las leyes de la genética de Mendel. Que la ley biogenética sea falsa ha sido el consenso de los biólogos durante más de 100 años, y los biólogos del desarrollo han estado trabajando de manera constructiva para ofrecer explicaciones alternativas, que hasta ahora todas han sido de naturaleza evolutiva.
- Las similitudes entre los embriones de vertebrados son reales. Debemos distinguir entre las observaciones de esas similitudes y las hipótesis sobre sus causas. Las similitudes no son objeto de duda; existen estudios valiosos sobre el grado y el momento de las similitudes, pero ninguno que cuestione su existencia general. Lo que Wells ha descrito es una hipótesis sobre la causa, la ley biogenética de Haeckel, que falló temprano y espectacularmente. No ha abordado ninguna hipótesis moderna, ni ha proporcionado una alternativa mejor.
- La evidencia para la descendencia común reside en la unidad de forma y proceso. No utilizamos el mecanismo anticuado e inválido de Haeckel para argumentar a favor de la evolución. En su lugar, observamos la maravillosa convergencia de organismos dispares en principios comunes: todos los animales utilizan los mismos genes para definir regiones de sus cuerpos, todos los vertebrados construyen sus caras mediante improbables reordenamientos de protuberancias faríngeas extrañas, e incluso los mamíferos sin cola como nosotros tenemos que comenzar con embriones con cola. La mejor explicación para estos fenómenos es que son una consecuencia de un patrimonio común.
[Volver a las preguntas frecuentes sobre los íconos de la evolución]