Resposta a Casey Luskin
por Nic Tamzek et al.[Última atualização: 14 de março de 2002]
![]()
Outros Links:
|
Introdução
A primeira versão do ensaio de Nick Tamzek, "Ícone da Obscuridade", era um panfleto bastante longo escrito para ser distribuído em uma palestra de Jonathan Wells na UCSD em 29 de janeiro de 2002. O texto deste panfleto foi publicado na web pouco depois, e várias versões de adições e edições foram publicadas sucessivamente até que o ensaio fosse publicado no talkorigins.org em 25 de fevereiro de 2002.
Logo em seguida, Casey Luskin do clube IDEA da UCSD IDEA club, um dos organizadores da palestra do Dr. Wells na UCSD, publicou uma resposta ao panfleto original (que ele está hospedando aqui) intitulado "Ícones Ainda em pé: Jonathan Wells sai limpo, apesar da crítica severa". O mesmo ensaio também já foi publicado no site do centro IDEA de San Diego IDEA center website e no site do ID Access Research Network (ARN).
Embora o folheto original carecesse de grande parte do material que se encontra no Arquivo TalkOrigins, a extensa resposta de Luskin tenta, de facto, abordar muitos argumentos chave do FAQ atual, e até agora é a única defesa conhecida de Wells. Assim, uma resposta a Luskin é merecedora. No entanto, recomenda-se que o Dr. Wells responda por si próprio.
Esta resposta foi montada pela técnica, em certa medida inovadora, de ter numerosos autores responder a trechos específicos do ensaio de Luskin. Uma das razões para isso é publicar uma versão até 11 de março, quando Wells e outros falarão em Ohio. Luskin apresentou uma lista tão longa de alegações criacionistas que uma única pessoa não consegue esclarecê-las todas em pouco tempo, mesmo que não sejam novas. Isso pode tornar a leitura um pouco desconexa, mas permite que várias pessoas contribuam em áreas de conhecimento relevante.
Para manter as coisas claras, as citações de Luskin estarão em caixas cinzas como esta. Todas as citações são copiadas diretamente da versão do ensaio de Luskin que foi salva da web em 3 de março de 2002. Portanto, alguns erros de digitação no original serão mantidos como estão, sem [sic], etc. A versão da defesa de Luskin sobre Wells que está postada no site ARN e no site IDEA está mudando constantemente conforme as pessoas apontam erros, então esta resposta, por necessidade, refere-se à versão de 3 de março. Além disso, note que este documento está em andamento. Parte do que Luskin escreveu será abordada em versões futuras deste documento.
Experimento de Miller-Urey
Abelson prossegue dizendo que não há "evidência para ... mas muito contra" uma atmosfera de amônia-metano, observando que "[u]ma quantidade de amônia equivalente ao nitrogênio atmosférico atual seria destruída [pela radiação ultravioleta] em ~30.000 anos."2 Ele, e Lasaga et al., descobriram que o gás metano teria deixado uma assinatura de carbono orgânico em rochas sedimentares antigas, talvez até mesmo devido a uma "mancha de óleo primordial" de 1 a 10 m de espessura15 (de qualquer forma, os experimentos de Miller-Urey produziram muito mais alcatrão do que qualquer outra coisa, e se muitos prebióticos foram produzidos, então o alcatrão deveria ter coberto a Terra primitiva), para o qual não há evidência geológica. Schopf observa que o Supergroupo de Swazilândia (~3,2 Ga) contém carbono inorgânico, "indicando que o dióxido de carbono, e não o metano, pode ter sido a forma dominante de carbono atmosférico neste ponto no tempo"16. Isso desafia a importância das fontes hidrotermais submarinas como contribuidoras de metano, o que Tamzek diz que Wells "exclui" incorretamente.
Como isso ocorre cerca de 300 milhões de anos após os primeiros fósseis bacterianos conhecidos e muito depois do período da origem da vida (4,2-3,8 Ga), alguém se surpreende? Além disso, no modelo de Kastings, o metano é uma forma minoritária (cerca de 20 ppm) e não dominante na atmosfera, tornando isso duplamente irrelevante para o modelo de fontes hidrotermais.
-- Ian Musgrave
A Árvore da Vida de Darwin
O Relógio de Darwin Está Ticking
Tamzek observa que a hipótese do relógio molecular está em grande disputa, o que é mais pronunciado quando o relógio molecular difere marcadamente do registro fóssil (especialmente em relação a mamíferos36, aves36 e grandes grupos animais36, 37). Os caracóis-cone79, que Tamzek alega citar como exemplo de evolução rápida, fornecem pouco material para refutar alegações criacionistas contra a origem natural da informação genética, pois isso apenas mostra que houve forte pressão seletiva sobre o mrRna do caracol-cone. Se a evolução ocorreu, nenhuma função nova foi criada além de alguns venenos destrutivos diferentes -- e esses venenos tiveram entre 20-50 Ma para desenvolverem suas formas bastante similares. O próprio artigo admite que seu exemplo mais marcante de suposta mudança rápida, que ele chama de "hipermutação focal"79, é "mecanicamente inexplicado", tornando explicações naturalistas, neste ponto, impotentes para explicar as supostas transformações genéticas.
Quando Luskin chega a abordar a duplicação gênica como um exemplo de evolução aumentando a informação, ele tem algumas coisas bizarras para dizer.
Este artigo, como muitos outros na literatura da biologia evolutiva, baseia-se na maior parte da evolução genética da bioquímica no mecanismo de duplicação gênica. No entanto, ao tentar evoluir algo, a explicação por duplicação gênica não ajuda muito no problema, porque, uma vez que você duplica um gene, você tem um novo pedaço de informação genética para brincar, mas o que isso te traz de bom? Se sistemas complexos precisam de partes específicas, que tipo de evidência existe de que esses genes duplicados serão as partes que você precisa?
É claro, duplicações gênicas ocorrem em organismos já vivos que não "precisam" de nada. No entanto, se uma duplicação produzir uma função adicional útil ou expandir uma função existente, então a seleção natural ajudará-a a se espalhar. Esta é uma das formas mais frequentes pelas quais novas informações funcionais são adicionadas aos genomas, e novos sistemas complexos são o resultado eventual de novos genes trabalhando juntos.
Luskin baseia-se em grande parte na sua rejeição em um artigo de Lynch e Conery [1], alegando que a taxa de duplicação de genes é muito baixa, mas o seu uso do artigo apresenta vários problemas sérios:
1. Lynch e Conery, ao utilizar genomas completos, tentaram derivar empiricamente a taxa de duplicações de genes únicos, fazendo isso excluindo famílias de genes múltiplos. Seu estudo não abordou a duplicação do genoma completo, duplicação parcial ou completa de cromossomos, ou outros tipos de duplicação segmental. Luskin rejeita a duplicação como um meio para gerar novidade biológica, mas ele falha em abordar os mecanismos mais potentes para a adição de material genético, os quais são capazes de adicionar centenas ou milhares de genes de uma vez.
2. Contrariamente à afirmação de Luskin de que a taxa de duplicação relatada é "bastante rara", ela é na verdade considerada bastante alta. Lynch e Conery afirmam que "50% de todos os genes em um genoma são esperados para se duplicar e aumentar em frequência alta pelo menos uma vez em escalas de tempo de 35 a 350 milhões de anos. Assim, mesmo na ausência de amplificação direta de genomas inteiros (poliploidização), a duplicação gênica tem o potencial de gerar um substrato molecular substancial para a origem de novidades evolutivas."
3. As metodologias e conclusões de Lynch e Conery foram ambas fortemente criticadas em um artigo de "comentários técnicos" da Science por dois conjuntos separados de cientistas [2], [3] que são especialistas bem respeitados. Entre outras coisas, a conclusão sobre a meia-vida curta de genes duplicados e a taxa de duplicação foram ambas questionadas, e embora Lynch e Conery defendam sua posição, eles admitem que várias suposições enviesam seus resultados e revisam algumas de suas estimativas. Portanto, os resultados do estudo são, no mínimo, altamente questionáveis. Luskin deveria ter mencionado essa controvérsia ou omitido o estudo por completo.
Até mesmo sem esses problemas, Luskin ainda falha em abordar adequadamente a questão da duplicação ao se basear neste estudo. A taxa apresentada por Lynch e Conery, mesmo que correta, é uma taxa média de duplicação. Mas diversos genes podem desviar-se muito da norma. Além disso, é bem conhecido que as taxas de duplicação gênica não são uniformes. Por exemplo, duplicações subsequentes são muito mais prováveis se uma duplicação já ocorreu anteriormente, porque as chances de crossing over desigual são muito aumentadas [4]. Isso permite a rápida expansão de famílias gênicas, o que parece ter ocorrido no caso dos venenos da caracol cone. Lynch e Conery deixaram as famílias gênicas de fora de seu estudo.
Lynch e Conery descobriram que a duplicação gênica média ocorre a cada 100 milhões de anos108--isso é bastante raro. Se as caracóis têm um tempo de geração de 1 ano, e o gene que você precisa se duplica uma vez a cada 100.000.000 de gerações, isso não lhe dá uma chance muito boa de obtê-lo quando você precisa. Além disso, descobriu-se que "a vasta maioria das duplicações gênicas é silenciada dentro de alguns milhões de anos, com os poucos sobreviventes posteriormente experimentando forte seleção purificadora"108. Outro estudo mostrou que não são muito livres para mutar por aí, que há forte pressão seletiva sobre eles107. Isso apoia a afirmação de Conery e Lynch de que os mecanismos reais pelos quais a duplicação gênica contribui para a evolução não são muito bem compreendidos:
"No entanto, não está claro como os genes duplicados navegam com sucesso uma trajetória evolutiva a partir de um estado inicial de redundância completa, onde uma cópia provavelmente é dispensável, até uma situação estável em que ambas as cópias são mantidas pela seleção natural. Nem está claro com que frequência esses eventos ocorrem."108
Luskin baseia-se em Lynch e Conery para afirmar que novos duplicados não duram muito tempo porque são rapidamente (ou seja, em milhões de anos) silenciados por mutação. (Ironicamente, os que ficam silenciados tornam-se pseudogenes, que oferecem evidências poderosas de descendência comum. Veja "Erros Plagiados e Genética Molecular", http://www.talkorigins.org/faqs/molgen/). Mas, além dos problemas mencionados anteriormente com aquele estudo, Luskin simplesmente ignora estudos recentes que mostram que duplicados são preservados muito mais frequentemente do que previamente pensado [5], [6] incluindo estudos por Lynch próprio [7], [8]. Além de evoluir funções completamente novas, duplicações gênicas podem particionar funções antigas que o gene ancestral executava apenas fracamente. Este modelo é apoiado pelo fato de que muitas enzimas são "promíscuas" em relação ao número de suas atividades fracas [9]. Isso permite que as funções particionadas sejam mais ativas e reguladas de forma diferente, às vezes sendo expressas em tecidos diferentes [10]. Desta forma, maior complexidade pode evoluir mesmo sem a adição de funções de novo, e duplicados são mais frequentemente preservados. Luskin conclui:
O ponto final é que a explicação por duplicação gênica ainda deixa os detalhes ao acaso, e esta via definitivamente não foi verificada experimentalmente. Tudo o que Espiritu et al. encontraram são homologias de proteínas, e então inferiram uma vaga via ancestral de criação gênica. Esta explicação para a origem de novidades evolutivas reais carece de um mecanismo confiável e é pouco melhor que uma explicação vaga.
Mas é claro que é Luskin quem está fazendo a "mão na boca". Ele se baseia em um estudo questionável que ele interpreta incorretamente, ignorando uma abundância de dados que contradizem sua tese. Contrariamente às suas alegações, os mecanismos para duplicação gênica, que incluem crossing over desigual e a inserção de retrogenes, são razoavelmente bem compreendidos [4] e são seguidos por qualquer número de possibilidades seletivas (veja qualquer uma das referências acima ou abaixo para informações extensas, ou qualquer uma das dezenas de outras). Além disso, a duplicação gênica adaptativa tem sido observada em laboratório [11], [12]. E observada entre espécies estreitamente relacionadas [13] (incluindo caracóis cone [14]) que até mesmo criacionistas da Terra jovem consideram relacionadas por descendência comum. Certamente, há mais coisas a serem aprendidas sobre duplicação gênica (o que é uma razão pela qual os venenos de caracóis cone são tão intensamente estudados), mas se ela ocorre ou não, ou sua importância na evolução, não está em dúvida. É simplesmente errado alegar que a duplicação gênica não é um mecanismo confiável para gerar novidade biológica.
REFERÊNCIAS
1. Lynch, M. e J.S. Conery, O destino evolutivo e as consequências de genes duplicados. Science, 2000. 290(5494): p. 1151-5.
2. Long, M. e K. Thornton, Duplicação gênica e evolução. Science, 2001. 293(5535): p. 1551.
3. Zhang, L., B.S. Gaut, e T.J. Vision, Duplicação gênica e evolução. Science, 2001. 293(5535): p. 1551.
4. Patthy, L., Evolução de Proteínas. 1ª ed. 1999: Blackwell Science Inc. 91-103.
5. Massingham, T., L.J. Davies, e P. Lio, Analisando a função gênica após a duplicação. Bioessays, 2001. 23(10): p. 873-6.
6. Dermitzakis, E.T. e A.G. Clark, Seleção diferencial após duplicação em genes do desenvolvimento mamífero. Mol Biol Evol, 2001. 18(4): p. 557-62.
7. Force, A., et al., Preservação de genes duplicados por mutações complementares e degenerativas. Genetics, 1999. 151(4): p. 1531- 45.
8. Lynch, M. e A. Force, A probabilidade de preservação de genes duplicados por subfuncionalização. Genética, 2000. 154(1): p. 459-73.
9. James, L.C. e D.S. Tawfik, Reatividades poliméricas catalíticas e de ligação compartilhadas por duas proteínas não relacionadas: O papel potencial da promiscuidade na evolução enzimática. Protein Sci, 2001. 10(12): p. 2600-7.
10. Serluca, F.C., et al., Partitioning of tissue expression accompanies multiple duplications of the Na+/K+ ATPase alpha subunit gene. Genome Res, 2001. 11(10): p. 1625-31.
11. Jelesko, J.G., et al., Rare crossing-over desigual germinativo levando à formação de genes recombinantes e duplicação gênica em Arabidopsis thaliana. Proc Natl Acad Sci U S A, 1999. 96(18): p. 10302-7.
12. Brown, C.J., K.M. Todd, e R.F. Rosenzweig, Múltiplas duplicações de genes de transporte de hexose de levedura em resposta à seleção em um ambiente limitado por glicose. Mol Biol Evol, 1998. 15(8): p. 931-42.
13. Cirera, S. e M. Aguade, Evolução molecular de uma duplicação: a região do gene do peptídeo sexual (Acp70A) de Drosophila subobscura e Drosophila madeirensis. Mol Biol Evol, 1998. 15(8): p. 988-96.
14. Espiritu, D.J., et al., Caracóis venenosos: filogenia molecular e a geração da diversidade de toxinas. Toxicon, 2001. 39(12): p. 1899-916.
-- Steve Reuland
Salvem as árvores!
Tamzek sugere que a árvore da vida de Darwin parece simples e "arvores" fora do 'matagal molecular' na raiz, no entanto, este não é o caso. É bem reconhecido na sistemática que, muito frequentemente, árvores filogenéticas baseadas em uma única sequência de gene ou proteína, levarão a uma árvore, enquanto uma árvore baseada em algum outro biomolécula parecerá bastante diferente. E ainda mais frequentemente, árvores baseadas em biomoléculas estão em desacordo com árvores criadas a partir de características macromorfológicas ou do registro fóssil (e árvores baseadas em diferentes macromorfologias frequentemente diferem umas das outras bem). Tamzek afirma que a árvore dos Eucariontes está bem resolvida através de Baldauf et al. (2000)99, no entanto, este artigo escapa dos problemas típicos de árvores conflitantes porque cria uma única árvore usando um conjunto de dados massivo de muitas sequências de proteínas para estatisticamente obscurecer as diferenças entre as árvores baseadas em proteínas individuais. Ao criar uma única árvore "frágil" através de muitos genes, a ancestralidade comum não pode ser fortemente verificada através de linhas convergentes independentes de evidência genética (reconhecendo, que esta é uma boa técnica para criar uma filogenia se se já assumir que a ancestralidade comum é verdadeira, no entanto, como um teste de ancestralidade comum, mostra que a evidência molecular fornece pouco suporte). De fato, foi encontrado que as filogenias de genes únicos apenas "apóiam subconjuntos da árvore de proteínas combinada." Em outras palavras, os diferentes genes, quando tomados independentemente, não convergem para fazer uma árvore bonita e organizada.
A alegação de Luskin de que Baldauf et al. combinaram os dados para obscurecer conflitos é infundada. Os autores reconheceram explicitamente o potencial de conflito entre os conjuntos de dados e, para abordar essa questão, analisaram as proteínas separadamente antes de combiná-las. Eis o que Baldauf et al. escreveram:
Análises de proteínas individuais mostram que nenhum agrupamento conflitante é apoiado por >65% aaBP e a maioria por muito menos (Fig. 3, colunas 1 a 4). Portanto, não existem conflitos fortemente apoiados entre esses dados. As análises par a par também mostram considerável evidência de cooperação filogenética entre todas as partições para a maioria das questões (Fig. 3, colunas 5 a 10); ou seja, a maioria das combinações par a par tende a apoiar as mesmas conclusões. Isso é mais uma evidência de histórias subjacentes comuns entre todas as partições. [Ênfase adicionada.]
Baldauf et al. não combinaram os dados para ocultar conflito porque não encontraram conflito significativo. Os clados que pareciam conflitar não tinham suporte estatístico e provavelmente foram resultado de dados insuficientes.
- Alex Wild
Tamzek afirma que os resultados deste artigo indicam que a filogenia dos Eucariotos está indo "bem", e que este artigo fornece uma filogenia dos Eucariotos "perfeitamente tradicional e em forma de árvore". Como dito anteriormente, a elegância desta árvore é puramente o resultado de técnicas estatísticas e do conjunto de dados combinado utilizado no artigo.
Parece que devemos adicionar a estatística à lista de disciplinas que devem ser falhas se os argumentos do Design Inteligente forem verdadeiros. A principal "técnica estatística" usada para produzir a "elegância" da árvore de Baldauf et al. é o uso de uma amostra maior de dados de caracteres. Se tirar de uma amostra maior for uma técnica estatística subastolada, porcos voam.
Seguindo a lógica de Luskin, deveríamos rejeitar o resultado de uma pesquisa de opinião com 100 pessoas em favor do resultado de uma pesquisa de opinião com 10 pessoas, porque a primeira utilizou uma "técnica estatística" que produziu um resultado diferente. Luskin nos levaria a pesar um resultado fracamente apoiado baseado em uma amostra pequena igualmente a um estudo muito mais robusto baseado em uma amostra maior.
Por que ele faz isso? Não sei. Acredito que o erro de amostragem produz resultados estranhos suficientes para que os defensores do design inteligente possam encontrar alguns para usar com fins retóricos, especialmente se eles desviarem das questões de tamanho da amostra e poder estatístico. No entanto, uma vez que começamos a pesar estudos não com base nos méritos de sua metodologia, mas por quão bem podemos usá-los em nossa retórica, deixamos de ser bons estudiosos e começamos a ser propagandistas.
-- Alex Wild
De qualquer forma, Baldauf et al. descobriram, na verdade, que sua árvore apresenta "diferenças marcantes da filogenia do rRNA SSU"99, e observaram que estudos anteriores também encontraram árvores conflitantes.
Vamos examinar o resumo do artigo de Baldauf et al., a fonte da citação:
A compreensão atual da sistemática de ordem superior dos eucariotos baseia-se em grande parte em análises do RNA da subunidade pequena ribossomal (SSU rRNA). O teste independente desses resultados ainda é limitado. Combinamos as sequências de quatro das proteínas mais amplamente amostradas taxonomicamente disponíveis para criar um conjunto de dados aproximadamente paralelo ao do SSU rRNA. A filogenia resultante mostra uma série de diferenças marcantes em relação à filogenia do SSU rRNA, incluindo forte suporte para a maioria dos grupos principais e vários supergrupos principais. [Ênfase adicionada.]
Observe, em particular, a frase que Luskin omitiu:
...incluindo forte apoio para a maioria dos principais grupos e vários supergrupos principais
O texto omitido altera significativamente o sentido da frase. Luskin estava fazendo o caso de que o rRNA do SSU fornece uma árvore diferente daquela produzida pelos 4 proteínas usadas por Baldauf et al., e em particular estava argumentando que os dois conjuntos de dados produzem árvores conflitantes. No entanto, no contexto, é claro que as diferenças entre o rRNA do SSU e as árvores de Baldauf et al. não estão principalmente na ordem de ramificação, mas no nível de suporte para os agrupamentos principais. Uma leitura adicional de Baldauf et al. confirma isso. A diferença entre forte suporte e fraco suporte para um agrupamento (ou de forte suporte para um agrupamento versus fraco suporte para um agrupamento diferente) é, com certeza, diferente da diferença entre dois agrupamentos fortemente suportados, mas conflitantes. O argumento de Luskin repousa na confluência dos dois.
Fonte: Baldauf SL, Roger AJ, Wenk-Siefert I, Doolittle WF. "Uma filogenia em nível de reino de eucariotos baseada em dados proteicos combinados." Science 290(5493):972-7 (3 de novembro de 2000).
-- Alex Wild
Embriões de Haeckel:
Developmental biologist PZ Myers was asked via email to comment on what Luskin wrote:
A Figura de Ampulheta Embriológica
Tamzek concede graciosamente que os desenhos de Haeckel não devem constar em livros didáticos (afirmando essencialmente o ícone), no entanto, ele cita Raff (2001)30 para sugerir que a embriologia ainda fornece evidências de uma história evolutiva compartilhada entre as classes de vertebrados. Raff sugere que Wells nega a "fase filotípica" e o "compartilhamento de elementos estruturais principais e seus elementos topológicos" entre embriões de vertebrados, no entanto Wells reconhece prontamente que as semelhanças embrionárias da "fase filotípica" realmente existem, dizendo: "aqui [na fase filotípica] as várias classes exibem pela primeira vez as características comuns a todos os vertebrados" (Ícones, p. 98). O ponto de Wells, aqui, é que os caminhos que os vertebrados em desenvolvimento percorrem para chegar à "fase filotípica" são muito diferentes, formando um padrão de desenvolvimento que o próprio Raff diz ter a forma de uma "ampulheta"32. Wells observa que este padrão de "ampulheta" não é previsto se todos os vertebrados compartilharem um ancestral comum.
Isso está correto. Um padrão específico de desenvolvimento não é uma previsão evolutiva; a ampulheta do desenvolvimento é uma observação. Observamos um período filotípico repetidamente em diferentes espécies, o que levanta uma pergunta interessante: por que os animais fazem isso? A explicação evolutiva é que existe uma base morfológica (o plano corporal) que é estabelecida em um período inicial do desenvolvimento, que tem sido refratária à mudança. Elaborações posteriores sobre o plano corporal baseiam-se nos padrões estabelecidos na fase filotípica, razão pela qual essa fase, amplamente definida, persiste.
Além disso, podemos fazer previsões com base em hipóteses sobre como os planos corporais são estabelecidos. Sabemos que os genes Hox são centrais para esse processo. Uma previsão que pode ser feita é que organismos com diferentes planos corporais devem apresentar mudanças correlacionadas na maneira como seus genes Hox são expressos, ou na maneira como os genes Hox regulam genes mais a jusante. Essas previsões foram testadas e estão atualmente sendo testadas em laboratórios ao redor do mundo.
O campo criacionista/design inteligente carece de qualquer explicação para as características comuns que observamos em organismos amplamente distintos. Se suas 'teorias' tivessem algum poder, deveriam ser capazes de nos ajudar a entender essas observações. Elas não são.
Myers:
[...]
Richardson afirma que isso não é um problema para a evolução darwiniana porque "a mistura de semelhanças e diferenças entre embriões de vertebrados reflete mudança evolutiva em mecanismos de desenvolvimento herdados de um ancestral comum". No entanto, ao examinar os dados que Wells apresenta em seu capítulo sobre Homologia em Apêndices de Vertebrados, descobrimos que os mecanismos que controlam o desenvolvimento muitas vezes não são previstos pela filogenia. Wells encontra que o gene Distal-less controla o desenvolvimento de apêndices no camundongo, verme espinhoso, borboleta, ouriço-do-mar e verme de veludo, mesmo que tais apêndices não tenham se originado através de uma ancestralidade comum. Não há razão pela qual a ancestralidade comum deveria prever que o mesmo gene produziria "pernas" em tantos tipos muito diferentes de organismos, quando seu suposto último ancestral comum não teria tido pernas.
Isso é parcialmente verdadeiro. É correto que não preveríamos que as pernas em artrópodes e vertebrados necessariamente teriam a mesma causa molecular, porque as pernas nessas duas linhagens não são homólogas — elas surgiram independentemente. No entanto, o que temos aqui novamente é uma observação. As pernas de vertebrados e artrópodes todas usam o mesmo gene, distal-less, para iniciar algumas das etapas iniciais do desenvolvimento dos membros! Esse é um resultado bastante surpreendente que favorece uma explicação evolutiva. Os evolucionistas sempre previram algum nível não especificado de similaridade entre diferentes linhagens, porque elas surgiram de um ancestral comum, mas não poderíamos saber o quão similares elas seriam até olharmos. Agora estamos na curiosa posição em que os criacionistas tentam argumentar que, porque as similaridades entre os organismos são mais fortes do que antecipávamos, isso é evidência contra a ideia de descendência comum!
Archaeopteryx: O elo perdido:
Vertebrate paleontologist Thomas R. Holtz, Jr. who showed just how inaccurate Luskin is. Holtz wrote in an email:
Fundamentado.
Tamzek afirma que os terópodes fósseis Dromaeosauridae (Protoarchaeopteryx e Caudipteryx) fornecem evidências inequívocas de que os pássaros evoluíram dos dinossauros.
Nem Protarchaeopteryx (observe a grafia) nem Caudipteryx são dromaeossáurios. São, respectivamente, um maniraptorano generalizado e um oviraptorossáurio.
No restante desta seção, as respostas a Luskin, em aspas, são do e-mail de Holtz. Ele forneceu o seguinte cladograma:

"Neste diagrama, os seguintes grupos possuem penas confirmadas/análogos de penas:
Avialae (aves) (óbvio...)
Dromaeosauridae (Microraptor,
Sinornithosaurus e a nova forma mencionada em
Nature nesta semana)
Alvarezsauridae (Shuvuuia)
Oviraptorosauria (Caudipteryx)
Therizinosauroidea (Beipiaosaurus)
Compsoganthidae (Sinosauropteryx)
Observe que, exceto Shuvuuia e as aves, todas as descobertas provêm de uma única Formação, o Yixian, notável por sua espetacular preservação. Observe também que não temos impressões de escamas ou quaisquer impressões não-penares para outras espécies nestes grupos particulares, portanto a explicação mais simples é que seu ancestral comum mais recente possuía penas simples."
"The dinosaurs from Tyrannosauroidea up to birds on this cladogram are noted by a reduction in the number of tail vertebrae; this is particularly true of birds and their closest relatives."
No entanto, o cético deve fazer a pergunta: isso é realmente o caso? Estes são fósseis interessantes, mas não fornecem evidências suficientes para colocar a hipótese dino-pássaro em prática. De fato, as caudas pesadas e os grandes corpos dos dinossauros terópodes são exatamente o oposto do tipo de organismo que esperaríamos evoluir para se tornar um voador92.
"These guys are mixed up; the description they refer to sounds like Sinosauropteryx (a compsognathid). No one has questioned the feathers on Protarchaeopteryx; the only question was whether or not it was a bird."
De acordo com o relatório em Nature38, o Protoarchaeopteryx era um dino com penas semelhantes a "pelos" no corpo, no entanto um paleontólogo chamou esse pelo de "dino-pelo, [que] realmente não poderia ter nada a ver com a origem das penas."91
De qualquer forma, as penas de voo não têm função para o voo nos pássaros. As únicas "penas com varetas, barbas e simétricas"38 em Protoarchaeopteryx aparecem em sua cauda — não há sinais de penas para o voo. Caudipteryx tem algumas remiges com varetas e barbas em um dedo da mão, mas seu braço é muito mais curto que o de um pássaro39 — muito curto para o voo. Como esses organismos possuem algumas estruturas corporais bem adaptadas ao voo, mas claramente não voaram, esses (e outros) fósseis levaram os evolucionistas a acreditar que algumas das estruturas complexas primárias especificadas para o voo — penas, asas e ossos ossificados — originaram-se para um propósito diferente do voo40.
Nenhum desses dois tem "asas"; são apenas braços. Os de Caudipteryx, na verdade, são realmente curtos em comparação com outros oviraptorossaurídeos!
"'ossos ossificados' = ossos feitos de ossos. Como você e eu temos ossos ossificados, assim como gatos, cães, lagartos, sapos e trutas, isso dificilmente é uma característica de voo!!"
Não há explicações elegantes aqui, mas sim pensamentos sonhadores tentando forçar os dados a se encaixarem em um modelo de evolução que falha em explicar a origem do voo.
"Na verdade, o que existe é a descrição do material em questão. Fóssis primeiro, pessoal; hipóteses segundo."
"Nope. These have been shown to be structurally disctinct from the true feathers of birds and other dinosaurs; see recent papers by Reisz and Sues."
Répteis de penas não se agrupam
De fato, penas também foram encontradas em um réptil semelhante a um lagarto não-dinossauro90,
no entanto, os defensores da hipótese do dinossauro-pássaro pensam que este fóssil emplumado está completamente desconectado da origem dos pássaros e dos supostos dinossauros emplumados. Então, por que a suposta aparência de penas nesses Dromaeosaurios pós-avianos deve se tornar a prova definitiva da ancestralidade dinossaúrica dos pássaros? Talvez esses "dinos emplumados" sejam apenas o que seus répteis escamosos
"Longisquama não é um squamate!!! (Squamate = lagartos & cobras). Também não é um verdadeiro archosaur, como tradicionalmente se considera.
"Além disso, a evidência primária da origem aviana dos dinossauros é esquelética, não integumentar. Consulte meu site, entre muitas outras referências."
"Identical."
os equivalentes são: quimeras de répteis não relacionadas a aves.
Para lançar mais alguns ossos na hipótese dino-ave, embora tanto os dinossauros terópodes quanto as aves andem sobre 2 pernas e possuam algumas semelhanças esqueléticas, diferenças como a configuração dos dígitos,
"identical (and in fact the same thing: pubis bone being part of the pelvis!) in basal dromaeosaurids, basal troodontids, and basal birds."
osso púbico, formato da bacia,
"Teeth of primitive birds, primitive troodontids, and primitive dromaeosaurids all very similar."
dentes,
"Fair to say that it is distinct from all LIVING reptiles."
e configuração de órgãos internos (o sistema respiratório aviano é único e muito diferente do dos répteis)
And rejected...
têm sido todas levantadas
as diferenças que desafiam a hipótese dino-pássaro43, 92. Wells também observa que os métodos cladísticos que estabeleceram essa suposta relação ignoram o registro fóssil,
"Nonsense. Como uma análise cladística de formas conhecidas apenas a partir de fósseis poderia ignorar o registro fóssil?
"(Suspeito que ele quis dizer 'registro estratigráfico', mas isso é outro assunto)"
"Guilty. :-) "
assume uma história evolutiva,
e ignore os principais problemas com as implicações de que os dinossauros evoluíram para voar a partir do chão.
"????? As análises filogenéticas são hipóteses de relações; são o quadro pelo qual outros estudos (como as origens do voo ou coisas semelhantes) podem ser interpretados, mas as próprias análises são APENAS as inter-relações."
O texto completo do e-mail de Holtz está aqui.
Mariposas Peppered e os Tentilhões de Darwin:
Basta com a minha retórica criacionista—Wells está certo?
Em relação às mariposas, Tamzek cita Majerus, que, em resposta a Wells, admite que a hipótese do melanismo está "descreditada", o que é de fato o que é ensinado nos livros didáticos, validando a crítica geral de Wells. Nesta resposta de 2000 a Wells, citada por Tamzek, Majerus afirma que a predação diferencial por aves continua sendo responsável pela mudança na coloração das mariposas. No entanto, o mesmo Majerus escreveu em 1987: "parece certo que a maioria dos B. betularia repousa onde está oculta.... [e] que as áreas expostas dos troncos das árvores não são um local de repouso importante para qualquer forma de B. betularia"87, o que excluiria a possibilidade de que a predação por aves seja um fator importante em sua seleção. Majerus, um especialista, parece estar se contradizendo aqui ou retirando suas alegações.
Na correspondência citada, Majerus não diz que a ideia de que aves que se alimentam de mariposas camufladas de forma diferencial resulta em melanismo industrial foi "descreditada", ele diz explicitamente que há evidências substanciais para isso. Ele disse que a apresentação nos livros didáticos é simplista demais, o que não valida a crítica de Wells.
Se o autor tivesse realmente lido a referência 87 (Howlett R.J., Majerus M., Biological Journal of the Linnean Society 30: 31-44 (1987)), ele teria notado que Majerus e Howlett não contestaram que a predação por aves era um fator importante na seleção naquele artigo. Sua preocupação era obter valores quantitativos precisos para a seleção via predação por aves. O autor também deveria ter notado que Howlett e Majerus relataram os resultados de um experimento comparando a pressão seletiva em locais expostos e não expostos. Não houve diferença significativa na proporção de mariposas escuras e claras capturadas por aves entre locais expostos e não expostos, portanto, a predação por aves é um fator importante em ambos os locais. A posição de Majerus em 1987 era a mesma de 2000.
-- Ian Musgrave
Independentemente do que as mariposas realmente façam, Wells não estava injustificado em confiar em Majerus, um especialista (e outros88), quando Majerus sugeriu que os pássaros não são um fator na seleção das mariposas87. Majerus e outros podem discordar de Wells, no entanto, Wells em si não é completamente ignorante sobre o assunto das mariposas, pois ele teve um artigo de opinião publicado sobre o tema intitulado, "Segundas Pensamentos sobre Mariposas Peppered"88 na revista revisada por pares, The Scientist.
A afirmação de revisão por pares é uma declaração falsa. The Scientist é descrito como "The News Journal for the Life Sciences" (http://www.the-scientist.com/). Em uma breve pesquisa no site, não consigo encontrar nada mencionando revisão por pares ou a falta dela, nem quaisquer diretrizes de submissão. Isso não é um bom sinal. De qualquer forma, o artigo de Wells estava na seção "Opinião", como você pode ver aqui.
Também enviei e-mail ao The Scientist para esclarecimentos sobre a revisão por pares. O editor, Alexander M. Grimwade Ph.D., respondeu:
1. THE SCIENTIST não é uma revista revisada por pares.
2. O artigo escrito por Wells foi publicado em nossa seção Opinião, onde permitimos que cientistas "se expressem" com edição mínima ou revisão do conteúdo. Para citar nosso masthead:
"THE SCIENTIST serve seus leitores de muitas maneiras, mas um de seus aspectos mais valiosos é seu compromisso com a discussão aberta de temas controversos. Embora os leitores frequentemente nos elogiem por esse compromisso, também devem reconhecer que todos os artigos publicados em THE SCIENTIST refletem as visões de seus autores e não as visões oficiais da publicação, sua equipe editorial ou sua propriedade."
O fato de que The Scientist pode não ter compreendido totalmente a situação das mariposas ao aceitar o artigo de Wells é indicado pela sua charge. Quem iria caçar mariposas com jaleco e com uma lupa?
Além disso, a versão original da resposta de Luskin dizia que o artigo de Wells em The Scientist "continha mais de 45 referências" – na verdade, ele tinha 17. A versão expandida do ensaio de Wells sobre a mariposa está disponível no ARN, o que provavelmente é a que Luskin estava se referindo, embora uma contagem cuidadosa da lista de referências revele 44 referências. De qualquer forma, o número de referências é irrelevante se elas forem usadas de forma injusta, como mostra a versão expandida da seção sobre a mariposa-do-pimenta do meu artigo.
Se, no entanto, você é um verdadeiro glotão de detalhes bem referenciados sobre traças-do-carvão e Wells, então a pessoa a consultar é Ian Musgrave. Alguns de seus comentários estão online aqui.
-- Nic Tamzek
Majerus nunca implicou que a predação por aves não era um fator importante na seleção. Na verdade, ele afirma explicitamente que é, e enfatiza que sua preocupação é obter uma estimativa quantitativa precisa da pressão de seleção devido à predação por aves. O fato de Wells ter sido publicado em "The Scientist" não diz nada sobre seu conhecimento de mariposas. "The Scientist" não é uma publicação revisada por pares, mas uma publicação dedicada a itens de interesse geral e notícias. Mais do que uma examinação superficial do artigo de Howlett e Majerus teria mostrado que Majerus não estava contestando que a predação por aves era um fator importante (embora não necessariamente exclusivo) no melanismo industrial. Não é que isso seja surpreendente, nem mesmo Kettlewell pensava que a predação era o único fator envolvido (Kettlewell, HBD. (1961) Ann. Rev. Ent., 6: 245-262.).
-- Ian Musgrave
Não para acumular aqui, mas este ponto não pode ser enfatizado o suficiente. Majerus repetiu e claramente expressou sua visão de que as histórias dos livros didáticos são excessivamente simplificadas, mas que o ponto básico, a saber, que a predação de mariposas por aves é a força seletiva principal causando a mudança na cor das mariposas, continua sendo uma boa conclusão bem sustentada pelos dados.
Aqui estão as conclusões de Majerus sobre o caso da mariposa-do-telha, ao final dos dois capítulos (quase sessenta páginas) de seu livro Melanismo Industrial que ele dedica precisamente a este tópico:
O caso da traça do mariposa é, sem dúvida, mais complexo e fascinante do que a maioria dos livros de biologia tem espaço para abordar. Mesmo aqui, tive de ser seletivo sobre quais conjuntos de dados incluir, dada a enorme quantidade de material disponível sobre este caso. Tenho tendido a concentrar-me no trabalho realizado no noroeste industrial da Inglaterra, porque este é o trabalho mais extenso, e na East Anglia, onde eu próprio trabalhei. O meu objetivo neste e no capítulo anterior tem sido fornecer informação suficiente para que o leitor avalie por si mesmo quais partes da história estão completas e onde é necessário trabalho adicional (ver Capítulo 10). A minha visão sobre o surgimento e declínio da traça do mariposa melânica é que a predação diferencial por aves em regiões mais ou menos poluídas, juntamente com a migração, são principalmente responsáveis, quase à exclusão de outros fatores. (Majerus, 1998, Melanismo Industrial, p. 155, ênfase adicionada)
-- Nic Tamzek
Do Macaco ao Humano: O Ícone Supremo:
Anthropologist Jeffrey K. McKee, author of A Cadeia Enredada: Acontecimento, Coincidência e Caos na Evolução Humana, provided a reply to this via email:
O paleontólogo Stephen Jay Gould provavelmente concordaria com Wells, uma vez que ele escreveu: "[m]ais da metade dos fósseis de hominídeos, embora sirvam de base para especulações intermináveis e elaboradas narrativas, são fragmentos de mandíbulas e pedaços de crânios."48 Roger Lewin também estaria de acordo quando escreveu: "o principal problema tem sido o lamentavelmente pequeno número de fósseis de hominídeos sobre os quais os prehistoriadores puderam exercer seus talentos interpretativos"75. Wells não é de forma alguma desonesto ao afirmar que a escassez de fósseis úteis para investigar a alegada ancestralidade hominídea dos humanos causa subjetividade entre os pesquisadores. As declarações de Wells estão em linha com o que Gould e Lewin disseram e não são negadas por um nível reduzido de relevância encontrado na citação de Henry Gee por Wells.
Se alguém está tentando estabelecer evidências de que os humanos evoluíram, então a verdade é que os paleoantropólogos têm um excesso de riquezas. Existem milhares de fósseis pelos quais documentar as principais tendências ao longo dos últimos 4 milhões de anos, começando com as primeiras criaturas bípedes (Australopithecus) que mantiveram membros superiores para escalar e que possuíam apenas um mínimo de expansão cerebral em comparação com os macacos vivos. O Homo inicial (ou seja, Homo habilis) mostrou aumentos adicionais no tamanho relativo do cérebro, e embora os membros tenham mudado sutilmente, suas proporções de membros permaneceram semelhantes às dos macacos. O Homo erectus/ergaster levou as tendências adiante, com hominídeos alcançando um tamanho corporal moderno e proporções corporais gerais, mas não a forma facial moderna ou o tamanho relativo do cérebro. O Homo sapiens arcaico então mostra uma mistura de características que apareceram em diferentes tempos e lugares, com um conjunto totalmente moderno de características aparecendo há cerca de 100.000 anos.
Se alguém está tentando decidir "quem gerou quem" e estabelecer uma filogenia precisa, ou deduzir as causas evolutivas imediatas para a origem e disseminação de características particulares, então mais fósseis ajudariam a resolver muitas das perguntas específicas ... é isso a que Gee, Lewin, Holden e Gould se referiam. Os cientistas querem discernir o "quem, quando, onde e como" da evolução, não apenas o "se".
McKee:
Faltando: Link
Do precário pouco que é encontrado, a conclusão final é que basicamente existem 2 tipos de fósseis de hominídeos: os do gênero Homo e os do gênero Australopithecus. Tudo do gênero Homo (Homo erectus, habilis, neanderthalensis, sapiens, etc.) possui crânios e formas corporais que são muito semelhantes aos humanos modernos—frequentemente até o ponto de que poderiam estar dentro da possível variação genética humana moderna.
Como notado acima, Homo habilis tinha proporções de membros semelhantes às dos símios, mas a morfologia de um bípede. Embora muitas das características individuais dos crânios de membros subsequentes do gênero Homo possam estar dentro da faixa humana moderna, o conjunto completo é notavelmente diferente. Por exemplo, muito se faz da observação de que a capacidade craniana de Homo erectus está dentro da faixa humana moderna. Bem, mas a proporção de tamanho do cérebro para tamanho do corpo está fora da faixa humana moderna.
McKee:
Tudo o mais do gênero Australopithecus (do qual o famoso fóssil "Lucy" é derivado), parece muito mais um chimpanzé (Australopithecus significa "macaco do sul"). Existem formas "robustas" de Australopithecus que não são muito semelhantes a chimpanzés, mas estas também parecem absolutamente nada com Homo, e os evolucionistas não acreditam que elas estejam na linhagem humana.
De fato, Australopithecus tem, em muitos aspectos, uma aparência semelhante à dos chimpanzés. Também possui um tamanho relativo do cérebro, dentes, ossos e postura e marcha inferidas, mais próximos dos humanos do que dos chimpanzés ... exatamente como se poderia prever para um membro da linhagem hominídea que evoluiu pouco depois da divergência de um ancestral comum das linhagens que levam aos humanos e aos chimpanzés modernos.
McKee:
Então, quais fósseis existem para ligar os membros semelhantes a chimpanzés de Australopithecus aos fósseis do gênero Homo, que basicamente representa os humanos modernos? A resposta é nada. Entre os dois existem grandes diferenças esqueléticas em relação à locomoção49, 51 e uma grande lacuna no tamanho do cérebro50, sem nada no meio. O "elo perdido", parece, ainda está perdido.
A natureza intermediária do esqueleto pós-craniano de Homo habilis, como mencionado acima, contradiz a afirmação anterior sobre a locomoção. Mas onde está a "grande lacuna" no tamanho do cérebro? Por exemplo, Australopithecus africanus tem uma capacidade craniana média de 440 cc (centímetros cúbicos), enquanto a média para Homo habilis é de 610 cc — e seus intervalos se sobrepõem. Não há um grande salto ali. Mais útil, no entanto, é uma medida do tamanho relativo do cérebro: o Quociente de Encefalização, ou EQ. Basicamente, o EQ nos diz quanto mais cérebro um animal tem em comparação com um mamífero típico do seu tamanho corporal. Assim, um chimpanzé, por exemplo, tem um EQ de 2,0 — o dobro do cérebro esperado para um mamífero do tamanho de um chimpanzé. Agora, vamos olhar para os EQs médios dos principais táxons fósseis de hominídeos e procurar grandes lacunas: Australopithecus afarensis: EQ=2,2; A. africanus: EQ=2,5 (consideravelmente mais do que um chimpanzé moderno, a propósito); Homo habilis/rudolfensis: EQ=3,1; Homo erectus inicial: EQ=3,3; Homo erectus tardio: EQ=4,0; Homo sapiens moderno: EQ=5,8.
O "elo perdido" ainda está perdido? Sim, pois toda vez que os paleoantropólogos encontram mais um elo fossilizado da cadeia, os criacionistas encontram dois elos mais perdidos (ou seja, entre o elo e seu antecessor, e entre o elo e seu descendente). Na taxa atual de descoberta e publicação de hominídeos fósseis, o número de elos perdidos deve aumentar em uma ordem de grandeza até 2050.
Após fornecer citações de várias autoridades, incluindo Gee, que destacam a falta de fósseis de hominídeos e a natureza subjetiva de muitas árvores filogenéticas da evolução humana, Wells está justificado em perguntar: "[g]iven the highly subjective nature of paleoanthropologoy--as acknowledged by its own practitioners--what can the field reliably tell us about human origins?" (Icons pg. 223). A resposta, parece, é definitivamente não que evoluímos sem dúvida de macacos.
[Nota: a seguinte resposta à seção "Missing: Link" de Luskin foi escrita antes da resposta de McKee ter sido fornecida.]
Luskin afirma que os fósseis de hominídeos podem ser divididos em dois grupos: os Australopithecus semelhantes a chimpanzés e os Homo, nos quais ele inclui o Homo habilis, dizendo que eles têm "crânios e formas corporais que são muito semelhantes aos humanos modernos". Isso é absurdamente errado. Embora seja verdade que os australopitecinos possuíam muitas características corporais semelhantes às de macacos, quantos chimpanzés caminham bípedes (sobre duas pernas) como faziam os australopitecinos? E embora as evidências pós-cranianas (ou seja, não parte do crânio) para o Homo habilis sejam escassas, é claro a partir das evidências fósseis (em particular OH 62) que ele tinha uma forma corporal semelhante à dos australopitecinos. Hoje, o que historicamente foi classificado como homo habilis, os habilines, é dividido em duas espécies, H. habilis e H. rudolfensis, por muitos pesquisadores. O H. habilis inclui um crânio que nenhum criacionista especial considera humano, enquanto o H. rudolfensis tem um crânio que alguns criacionistas pensam ser de um macaco e outros pensam ser de um humano, e ainda assim os dois são muito semelhantes. Recentemente, Bernard Wood, um pesquisador influente, e Mark Collard publicaram "The Human Genus" em Science (vol. 284, pp. 65-71, 2 de abril de 1999), no qual propuseram que ambas essas duas espécies habiline sejam reclassificadas como Australopithecus e não como Homo. Vários outros pesquisadores já propuseram anteriormente que alguns dos fósseis habiline deveriam ser considerados australopitecinos. E Luskin quer que acreditemos que há uma linha clara entre Homo e Australopithecus? Mas deixarei que os leitores julguem por si mesmos. A tabela a seguir tem algumas imagens de alguns crânios de Homo ergaster que tinham uma forma corporal bastante moderna, as duas espécies de habilines e uma espécie de australopitecino. Os links abaixo das imagens levam a uma foto maior ou a mais informações sobre o fóssil em questão. As capacidades cranianas estimadas (tamanhos dos cérebros) de cada espécie foram retiradas do artigo de Wood e Collard.
| Espécie | Capacidade Craniana |
FH = Perguntas Frequentes sobre Hominídeos Fósseis SI = Smithsonian Institution Human Origins Program |
||
|---|---|---|---|---|
| Homo Ergaster | 854 cm3 |  FH SI |
 FH SI |
|
| habilines | Homo (or Australopithecus) rudolfensis | 752 cm3 | 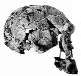 FH SI |
|
| Homo (or Australopithecus) habilis | 552 cm3 | 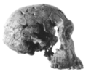 FH SI |
 FH SI |
|
| Australopithecus africanus | 457 cm3 |  FH SI |
 SI |
|
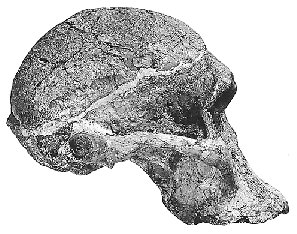{kind=link}
Há muitos outros fósseis também. O leitor é convidado a ir para este link e tentar traçar a linha entre "primata" e "humano." Não é uma questão simples como se poderia esperar se primatas e humanos fossem criações separadas, como demonstrou a tabela de opiniões de criacionistas de Jim Foley sobre o que são os hominídeos fósseis.
-- Mike Hopkins
Conclusão:
Wells não é um fraudador
Além de suas dúvidas sobre o darwinismo, uma revisão das acusações de Tamzek contra Wells revela que não há um único caso estabelecido em que Wells esteja errado, e poucos em que ele esteja claramente em desacordo com o pensamento científico mainstream. O único erro bona-fide em Icons poderia ser encontrado na figura 5-3 de Wells sobre a explosão cambriana, que Tamzek não verifica, e este suposto erro em si não afetaria a força dos argumentos de Wells sobre a explosão cambriana. Como é que Wells pode parecer estar em acordo com a ciência mainstream na quase totalidade de seus argumentos (mesmo Eugenie Scott diz que Wells é "tecnicamente correto" na maioria das vezes89), o que deveria ter um efeito profundamente negativo sobre as evidências para a teoria da evolução, e ainda assim Wells difere da ciência mainstream em sua conclusão de que a evolução pode ser falsa? A resposta mais provável reside na interpretação de Wells dos dados que coleta ao analisar o nível de erros nos livros didáticos.
O texto completo da resenha de Eugenie Scott pode ser encontrado em:
http://www.scienceormyth.org/icons%20of%20evolution.html
Luskin cita-a fora de contexto, como foi apontado aqui. Aqui está um pouco mais do que ela escreveu:
...Isso ocorre porque Wells apresenta uma visão sistematicamente enganosa sobre a evolução. As frases individuais em Icons são geralmente tecnicamente corretas, mas são artfully conectadas para levar o leitor fora do caminho da verdadeira biologia evolutiva e em direção a um matagal de mal-entendidos.... [Ênfase adicionada.]
-- Mike Hopkins
Dê um passo à luz
Em conclusão, Tamzek cita Teodosio Dobzhansky, um evolucionista teísta, para mostrar que se pode crer em Deus e na evolução. Na citação que Tamzek fornece, Dobzhansky parece estar argumentando apenas contra uma perspectiva de Terra jovem, o que não é nada do que Wells defende. Na verdade, Wells poderia até concordar com Dobzhansky de que os textos religiosos não são "livros-texto de astronomia, geologia, biologia e antropologia". Onde Wells discordaria é que a conclusão, portanto, deveria ser que, empiricamente falando, Deus não teve efeito direto sobre o processo de criação, e que Deus criou tudo através da evolução não direcionada. A Escritura Bíblica pode dizer pouco sobre os detalhes dos processos reais que Deus usou para criar, no entanto, está claro que Deus pretendia que as coisas fossem como são57. Dobzhansky teria dificuldade em encaixar a aleatoriedade da evolução na propósito de Deus. Para o teísta disposto a arriscar sua credibilidade acadêmica reconhecendo contraprovas à vaca sagrada da biologia105, esses problemas podem ser resolvidos. Se Dobzhansky tivesse vivido para ver o nascimento do movimento de design inteligente, talvez ele também, como teísta, teria abraçado sua abordagem mais aberta.
Em algum nível fundamental, todos os criacionistas acreditam, em geral, no design inteligente. Isso significa que, seja como Ele escolheu fazê-lo, todos os criacionistas acreditam que Deus criou o mundo e suas espécies, incluindo o homem, de forma intencional. Em seu ensaio, "Nada na Biologia Faz Sentido Exceto à Luz da Evolução", Dobzhansky explicitamente se autodenominou criacionista e evolucionista, o que, por substituição, significa que ele se considerava um defensor do design inteligente evolutivo ou da evolução teísta. (A partir de agora, todas as referências a "design inteligente" serão destinadas a referir-se ao "design inteligente anti-evolucionista" e não à evolução teísta. No entanto, deve-se enfatizar que, em geral, não há necessidade de uma inteligência utilizar processos estritamente não-darwinianos para criar as diversas formas de vida neste mundo. A evolução teísta é uma posição legítima do design inteligente e, notavelmente, é apenas a versão de Wells ou de sua espécie do design inteligente anti-evolucionista que faz da evolução ou dos métodos da ciência um inimigo.) Portanto, tentar apoiar a ideia de que Dobzhansky defenderia qualquer forma de criacionismo anti-evolucionista, especialmente as formulações modernas do design inteligente, seria um grande desafio.
Em sua resposta a Tamzek, Luskin tenta realizar precisamente essa façanha hercúlea. Luskin tenta argumentar que Dobzhansky era ignorante tanto das contribuições da sorte aleatória para a evolução quanto dos métodos modernos de design inteligente para explicar observações. Se verdadeiro, Luskin raciocina que Dobzhansky poderia ter apoiado o raciocínio moderno de design inteligente se simplesmente tivesse sido dado a chance. Infelizmente, essas afirmações não são verdadeiras e a alegação de Luskin de possível apoio de Dobzhansky é improvável.
Primeiro, o movimento do design inteligente não é novo. Suas raízes modernas remontam, pelo menos, a Michael Polanyi, que foi contemporâneo de Dobzhansky. Mesmo omitindo as contribuições modernas de Polanyi à literatura do design inteligente, o "Argumento do Design" do Reverendo William Paley descreveu pela primeira vez o argumento básico do design inteligente, e Paley viveu muito antes mesmo de Darwin.
Em um nível básico, os argumentos de design inteligente afirmam que um certo fenômeno não pode ser explicado pela ciência e, portanto, necessitam da existência de Deus. [2] As contribuições de Behe e Dembski para a literatura de design inteligente, modernas e esotéricas como podem parecer, ainda se baseiam na mesma lógica falha de Paley; apenas o fenômeno em questão mudou ao longo do tempo. Portanto, os argumentos de design inteligente não mudaram de forma significativa desde que Paley os articulou pela primeira vez. Como Dobzhansky não apoiou o raciocínio de Paley, ele provavelmente não teria apoiado o raciocínio de Behe ou Dembski também.
Existe também uma alta probabilidade de que os argumentos modernos de design inteligente sejam projetados para contornar a cláusula de estabelecimento de nossa Constituição. De fato, os defensores modernos de design inteligente removeram apenas as afirmações positivas sobre o projetista que Paley fez: onde Paley argumentava explicitamente por Deus, os defensores modernos de design inteligente argumentam por um "projetista" mais vago e oficialmente não nomeado. Tal manobra retórica não altera realmente a lógica do argumento de Paley. Pelo contrário, ela simplesmente torna incerta a identidade do projetista, os propósitos do design do projetista e outras afirmações positivas sobre o design. Novamente, o argumento dos defensores modernos de design inteligente não é substancialmente diferente do argumento dos defensores de design inteligente pré-modernos.
Para Luskin alegar que Dobzhansky nunca teve a oportunidade de considerar métodos ou argumentos de design inteligente é negar tanto a longa história do criacionismo do design inteligente quanto a congruência entre os argumentos modernos e pré-modernos de design inteligente. Em vez disso, é provável que Dobzhansky tenha previsto a inclusão deles mesmo enquanto fazia o caso contra os criacionismos anti-evolucionários em seu ensaio que antecedeu tanto Behe quanto Dembski.
Em segundo lugar, a evolução envolve, pelo menos em parte, uma contribuição da sorte aleatória — sempre envolveu, desde que Darwin escreveu sua primeira edição de Origem. Assim, como um cientista contribuinte para o campo da biologia e o estudo da evolução, é improvável que Dobzhansky não tenha considerado a aleatoriedade na evolução que ele pensava ser perfeitamente compatível com a teologia moderna. Considere a seguinte citação de seu ensaio.
A diversidade orgânica torna-se, no entanto, razoável e compreensível se o Criador criou o mundo vivo não por capricho, mas por evolução impulsionada pela seleção natural. É errado considerar a criação e a evolução como alternativas mutuamente exclusivas. Eu sou um criacionista e um evolucionista. A evolução é o método de criação de Deus, ou da Natureza. A criação não é um evento que ocorreu em 4004 a.C.; é um processo que começou há cerca de 10 bilhões de anos e ainda está em curso.
Uma seção inteira deste ensaio é dedicada à discussão da diversidade orgânica. Não há dúvida de que Dobzhansky atribuiu a aleatoriedade parte dessa diversidade. No entanto, as palavras "aleatório" ou "acaso" não aparecem no ensaio de Dobzhansky. Ainda assim, ele é bastante claro em outras obras de que a evolução envolve contribuições da aleatoriedade e do acaso. Considere esta citação do livro-texto de Futuyma:
Como disse Dobzhansky (1970), "Pode parecer uma imperfeição lamentável da natureza que a mutabilidade não seja restrita a mudanças que aumentem a adaptabilidade dos seus portadores. No entanto, apenas um vitalista Pangloss poderia imaginar que os genes sabem como e quando é bom para eles mutar." {Futuyma, Biologia Evolutiva 2ª ed. 1986, p. 76}
Ou considere esta citação do American Scientist:
O processo de mutação é a única fonte conhecida de novos materiais de variabilidade genética e, portanto, da evolução. {T. Dobzhansky, American Scientist, v. 45, 1957, p.385}
À luz dessas afirmações, parece que Dobzhansky reconheceu, de fato, o acaso em sua compreensão da evolução. "Nada na Biologia Faz Sentido Exceto à Luz da Evolução" foi escrito em 1973 e as citações acima foram escritas em 1970 e 1957. Por qualquer avaliação razoável, Dobzhansky incluiu influências aleatórias na evolução que acreditava ser compatível com o teísmo.
Luskin sugere que Dobzhansky era ignorante quanto à aleatoriedade na evolução, embora Dobzhansky soubesse muito bem da aleatoriedade envolvida na evolução. Luskin argumenta que Dobzhansky era ignorante quanto aos argumentos modernos de design inteligente, embora Dobzhansky soubesse muito bem das falhas na versão prototípica de Paley do mesmo. A sugestão de Luskin de que Dobzhansky poderia ter concordado com o criacionismo do design inteligente é, portanto, não suportada e, de fato, altamente improvável.
Ao contrário de Tamzek, Wells usa as citações de Dobzhansky para fins puramente científicos — para mostrar que a famosa citação de Dobzhansky — "[n]ada na biologia faz sentido fora da luz da evolução"104 — representa um afastamento do método científico, pois presume que tudo na biologia deve se encaixar no paradigma evolutivo:
"[C]iência em seu melhor estado busca a verdade. Dobzhansky estava completamente errado, e assim também estão aqueles que continuam a repetir seu mantra anticientífico. Para um verdadeiro cientista, nada na biologia faz sentido fora da luz das evidências." (Ícones, p. 248)
Dobzhansky escreveu um ensaio intitulado "Nada na Biologia Tem Sentido, Exceto à Luz da Evolução". Notavelmente, Dobzhansky escreveu um ensaio com este título; ele não escreveu apenas o título. No ensaio, Dobzhansky deixa claro por que escolheu este título para seu trabalho: a evolução explica as observações melhor do que qualquer outra explicação oferecida e a biologia sem evolução seria um amontoado de observações sem um tema unificador.
Considere os seguintes trechos do ensaio em questão:
O ambiente não impõe mudanças evolutivas aos seus habitantes, como postulado pelas agora abandonadas teorias neolamarckianas.
Tudo isso é compreensível à luz da teoria da evolução; mas que operação sem sentido teria sido, por parte de Deus, fabricar uma multitude de espécies ex nihilo e depois deixar que a maioria delas se extinguisse!
Elas imaginam que todas as espécies existentes foram geradas por um decreto sobrenatural há alguns milhares de anos, praticamente como as encontramos hoje. Mas qual é o sentido de ter de 2 a 3 milhões de espécies vivendo na Terra?
O Criador estava de humor brincalhão quando criou Psilopa petrolei para os campos de petróleo da Califórnia e espécies de Drosophila para viver exclusivamente em algumas partes do corpo de certos caranguejos terrestres em apenas algumas ilhas no Caribe?
Mas e se não houvesse evolução e cada uma das milhões de espécies tivesse sido criada por um ato separado de vontade? No entanto ofensiva que a noção possa ser para o sentimento religioso e para a razão, os antievolutionistas devem novamente acusar o Criador de trapacear. Eles devem insistir que Ele deliberadamente organizou as coisas exatamente como se seu método de criação fosse a evolução, intencionalmente para enganar sinceros buscadores da verdade.
Observe como Dobzhansky demonstra especificamente como considerou outras explicações, tanto não naturais quanto naturais, mas encontrou todas elas insuficientes. Após considerar essas alternativas, que provavelmente são representativas de todas as formas de criacionismo anti-evolucionário ou evolução não neodarwiniana (evolução lamarckiana, etc.), Dobzhansky intitulou seu ensaio como fez. Se Dobzhansky não tivesse considerado nenhuma alternativa, a objeção de Luskin seria precisa: há uma chance de que outra hipótese de biodiversificação se encaixasse nos dados; portanto, considerar a evolução como a única explicação possível é ou totalmente inválido ou dogmático.
Infelizmente para Luskin, o ensaio deixa bem claro que o título não transmite ou desenvolve plenamente as ideias de Dobzhansky. Pode ser possível explicar dados a partir de uma perspectiva não-evolucionista, mas, como Dobzhansky aponta frequentemente no ensaio, essas perspectivas são absurdas e não são tão sólidas ou úteis quanto a evolução. Assim, o título de Dobzhansky permanece válido e o ponto declarado por Luskin é facilmente descartado.
Existe outra maneira pela qual a objeção de Luskin poderia ser válida, embora ele não a formuleie por si mesmo. E se a evolução fosse facilmente removida do campo da biologia en bloc? Se os cientistas conseguissem, de alguma forma, compreender a biologia sem a evolução, então a sugestão implícita no título literal de Dobzhansky — de que nada na biologia pode fazer sentido sem a evolução — seria imprecisa e a objeção de Luskin poderia ser sustentada.
Mais provavelmente, foi a esta mesma objeção que Dobzhansky escreveu em seu ensaio:
Visto à luz da evolução, a biologia é, talvez, a ciência intelectualmente mais satisfatória e inspiradora. Sem essa luz, torna-se um amontoado de fatos diversos, alguns interessantes ou curiosos, mas que não formam uma imagem significativa como um todo.
Dobzhansky está dizendo que a separação da evolução de nossa compreensão moderna da biologia seria ou um processo difícil ou tornaria a biologia inteiramente incompreensível. Em qualquer outra linguagem, ele está dizendo que nada na biologia faz sentido exceto na "luz" organizadora e explicativa da evolução, uma interpretação justificável que também enfraquece a objeção de Luskin.
Criacionistas frequentemente tentam minar a lógica do ensaio de Dobzhansky alegando que existem, de fato, observações em biologia que não requerem uma perspectiva evolutiva para serem compreendidas. Wells parece seguir o mesmo caminho ao também argumentar contra o título em vez do ensaio, embora prefira atacar desmerecendo a evolução como mero paradigma. Ambos esses argumentos presumem que o leitor leu apenas o título e não o ensaio que o carrega. Se alguém lê o ensaio de Dobzhansky, a escolha do título é justificada, mesmo que seja um resumo truncado do próprio ensaio. Se Luskin está argumentando que o título do ensaio de Dobzhansky não desenvolve o ensaio de Dobzhansky tão completamente quanto o próprio ensaio faz, seu argumento é sem relevo e facilmente descartável ao lembrar aos leitores que a compreensão de questões raramente é alcançada ao julgar livros proverbialmente pela sua capa.
Na verdade, os pontos de Luskin e Wells são ambos facilmente abordados lendo o ensaio em questão [http://www.2think.org/dobzhansky.shtml]. O título desse ensaio é completamente justificado pelo desenvolvimento do ensaio: não é uma desviação do método científico, nem é defeituoso em seu raciocínio.
Quanto aos métodos da ciência, que ironia que Wells, como criacionista do design inteligente, acusasse Dobzhansky de ter se desviado do método científico. As respostas às perguntas biológicas que os defensores do design inteligente apresentam, porque essas respostas invocam "sobrenaturais" inobserváveis e imensuráveis como agentes de mudança natural, são respostas que não podem ser testadas pela ciência. São os criacionistas do design inteligente, e não Dobzhansky, que se desviam do método científico — e eles se desviam dele de forma ampla. Portanto, Wells não pode seriamente acusar ninguém de se desviar dos métodos da ciência sem envolver-se em flagrante hipocrisia.
O ponto de Wells é que a evidência — não um paradigma — deve controlar nossa compreensão dos dados. Após examinar as fraquezas nas linhas de evidência mais comuns usadas para apoiar a teoria da evolução, pelo menos duas coisas ficam claras: a evidência para a teoria da evolução certamente é insuficiente em alguns lugares importantes, e Ícones da Evolução será uma grande dor de cabeça para os evolucionistas nos próximos anos.
A compreensão científica atual do mundo "natural" observável e mensurável é aquela que explica as observações e prevê com maior precisão novos dados. Novos dados e novas teorias são considerados à luz dessas compreensões atuais. É o processo da ciência reavaliar continuamente o status das explicações consideradas, mantendo as teorias que melhor explicam e preveem os dados e desvalorizando ou rejeitando completamente o resto. Ao definir a evolução como a compreensão científica atual da biodiversificação, deve-se inferir que ela, portanto, explica e prevê melhor as descobertas vindas do mundo testável.
Wells reconhece que a evolução possui uma estrutura teórica ampla à qual novos dados são aplicados. No entanto, ele a condena como meramente um "paradigma", como se a evolução fosse uma teoria que, em princípio, não seria capaz de ser refutada por observações naturais. Ao fazer este argumento, ele ignora o fato de que, desde o dia em que Darwin concebeu a ideia, a evolução era capaz de ser refutada. Até o momento, isso não ocorreu.
Não é um tratamento tendencioso para a evolução ou contra o design inteligente que mantém a evolução como o entendimento científico atual para a biodiversidade e outras observações. Muito mais do que simplesmente um "paradigma" para guiar o pensamento em biologia ou não, a evolução é, em vez disso, uma peça fundamental de uma teoria científica. Ela guiou inúmeros experimentos, explicou tantas observações e medições em diversos campos e foi verificada e reforçada tão completamente que está tão próxima de um fato quanto a ciência provavelmente jamais chegará. É verdadeiramente "A teoria integradora da biologia."
Portanto, a sugestão de Wells de que a evolução é simplesmente um "paradigma" favorecido ou não possui um arcabouço teórico útil é completamente equivocada. A evolução é, em vez disso, a compreensão científica atual porque as evidências a suportaram robustamente como tal.
Quanto às evidências, Wells frequentemente se apresentou como um biólogo que simplesmente discorda da evolução. Se Wells realmente agisse como um cientista ou utilizasse efetivamente o treinamento que adquiriu ao buscar seu mestrado em biologia avançada, ele estaria trabalhando para desenvolver as evidências necessárias para sustentar seu caso contra a evolução. Em vez disso, Wells argumenta contra a definição de ciência, chamando a restrição da ciência a hipóteses testáveis e "naturais" de tendenciosa. Wells provavelmente reconhece a natureza robusta da evolução e, com o objetivo de promover sua admitida cruzada contra o darwinismo, para a qual buscou credenciais avançadas como biólogo em primeiro lugar, considera mais fácil minar os métodos da ciência do que sustentar cientificamente sua hipótese.
Em vez de sucumbir ao estratagema de Wells, o leitor é convidado a examinar as evidências que sustentam tanto a evolução quanto o design inteligente e tomar uma decisão como faria um cientista. A revista Evolução publica mais de 18 artigos revisados por pares rigorosamente a cada mês. Em todos os periódicos, vários artigos originais de pesquisa sobre evolução são publicados todos os dias, e inúmeros outros estudos são tornados possíveis apenas graças às insights que a evolução fornece ou às previsões que ela faz. Por outro lado, é realmente notável que o design inteligente não tenha nenhum desses artigos. Eles nem sequer tentam fazer pesquisa real porque não podem; nenhum dado jamais pode contradizer um agente sobrenatural postulado e apelar para as ações não observáveis e imensuráveis de agentes sobrenaturais para explicar dados naturais é simplesmente uma hipótese não falsificável na ciência.
Apesar da completa ausência de evidência cientificamente significativa, Wells e os outros criacionistas do design inteligente solicitam que a "evidência" para o design inteligente seja admitida ao lado da evolução nas salas de aula de ciências, com base na ideia de que a evidência, e não um "paradigma", deve controlar nossa compreensão dos dados. Se se baseasse realmente na evidência para a inclusão ou rejeição de discussões em salas de aula de ciências, o design inteligente simplesmente não apareceria em uma sala de aula de ciências. O argumento de Wells torna-se, portanto, tão hipócrita como o anterior: ele não tem evidência para sustentar sua alegação, a não ser um apelo à ignorância.
Luskin, invocando a pesquisa de Wells, afirma então que existem "falhas" na teoria da evolução. Sua descrição pejorativa da pesquisa atual como "falhas" é certamente digna de reprovação, que será apresentada. Primeiro, esta resposta deve começar lembrando aos leitores o conteúdo da crítica original de Tamzek a Wells. Qualquer investigador imparcial reconhecerá que Wells não pode ser confiável para transmitir adequadamente o estado da evolução aos leitores. Apenas porque Wells considera certas linhas de evidência em apoio à evolução como "flawed" não significa que observadores abertos as considerariam falhas. Luskin faria bem em encontrar outros apoios para suas conclusões do que as declarações no livro de Wells.
É lamentável, mas revelador, que Luskin descreva pejorativamente áreas ativas de pesquisa em evolução como "defeitos". Ao fazê-lo, ele intencionalmente sugere a ilegitimidade da evolução, mesmo enquanto a evolução atravessa uma gloriosa era de sucesso e utilidade. De fato, o uso pejorativo de Luskin revela mais sobre seu viés pessoal contra a evolução do que sobre a própria evolução. Claramente, há necessidade de mais pesquisa no campo da evolução; ninguém duvida disso. No entanto, mais pesquisa é necessária em evolução pelos mesmos motivos pelos quais mais pesquisa é necessária em química. Ou seja, sempre haverá mais a aprender sobre química e algumas das coisas que achamos que sabemos sobre química podem resultar-se erradas; no entanto, isso não apoia a alquimia.
O fato é que a ciência é uma disciplina ativa e há e sempre haverá uma fronteira além da qual ainda não temos compreensão e na qual nossa compreensão está incompleta. Nesta e nas proximidades desta fronteira, nossos modelos estão sujeitos a modificações, talvez até rejeição, conforme novas informações se tornam disponíveis. No entanto, a existência de incerteza nesta fronteira não nega nosso conhecimento no terreno bem trilhado do primeiro plano. [1] Com os devidos agradecimentos a Einstein, "Se soubéssemos tudo sobre [evolução], não seria ciência."
É verdade que ainda há muita pesquisa necessária sobre a evolução: os detalhes sobre os mecanismos da evolução, os caminhos específicos que ela tomou em linhagens particulares, a aplicação da evolução a outros problemas ou dados não necessariamente relacionados à especiação, etc., todos precisam ser investigados e resolvidos. No entanto, a intenção da declaração de Luskin — de que "defeitos importantes" existem na teoria da evolução e que esses "defeitos" minam a evolução como um todo — é inválida por pelo menos duas razões.
Primeiro, é importante entender que, no "cenário bem trilhado" dos fundamentos da evolução, ela está solidamente articulada com um apoio científico adequado — digamos, esmagador. O debate sobre a evolução ou as pesquisas realizadas sobre a evolução não é sobre se ela ocorreu, mas sobre como ocorreu. Colocado em outras palavras, no nível de ensino em que os conceitos evolutivos estão sendo ativamente investigados, provavelmente você estaria matriculado em um curso de pós-graduação: a evolução no nível do ensino médio e de graduação está tão próxima da factualidade quanto a ciência pode provavelmente chegar. Simplesmente, as áreas de pesquisa ativa na evolução — as supostas "falhas" de Luskin — não são do tipo que faria o imparcial duvidar de que a evolução ocorreu; as "fronteiras" da evolução estão muito distantes dos fundamentos estabelecidos, sobre os quais não há objeção científica séria. Portanto, Luskin está bastante errado ao alegar que há "falhas" na evolução, se sua intenção ao citar essas "falhas" é sugerir que elas minam a evolução.
Em segundo lugar, observe a pressuposição não declarada de Luskin: o fracasso da nossa compreensão atual da evolução pode ser tomado como um sucesso para o design inteligente. Essa lógica sugere que o design inteligente é uma explicação padrão: se descartarmos a evolução, o design inteligente é tudo que resta. Sua pressuposição é seriamente falha por pelo menos duas razões. Primeiro, uma falha comum de um argumento da ignorância é que superar a ignorância em questão facilmente mina o próprio argumento. De modo geral, dada a história da ciência e sua gloriosa história de superar a ignorância sobre fenômenos, as lacunas na nossa compreensão atual da evolução serão preenchidas eventualmente. Especificamente, a tentativa de Luskin de descartar as compreensões atuais da evolução como "falhas" não contribui para descartar compreensões futuras da evolução.
Em segundo lugar, e mais importante, o que há no design inteligente que faz Luskin pensar que ele merece o status padrão? O entendimento da evolução que temos atualmente — que possui áreas ativas de investigação em conceitos evolutivos avançados — tem muito mais um quadro teórico, preditivo, explicativo e útil do que o criacionismo do design inteligente, que francamente não possui nenhuma dessas coisas. Considerar que "a evidência para a teoria evolutiva certamente falta em alguns lugares importantes" e concluir que o design inteligente é, portanto, uma melhor explicação para observações perfeitamente naturais é abandonar a perspectiva, a razão ou os métodos da ciência.
Quanto ao livro de Wells ser uma "dor no cu para os evolucionistas", de fato ele assumiu o papel de um gadfly socrático, despertando respostas das comunidades científica e religiosa adormecidas. Anteriormente, essas comunidades não consideravam o criacionismo uma ameaça. Após a publicação do livro de Wells, elas tornaram-se mais ativas em abordar o criacionismo.
Como resultado, em qualquer local onde Wells ou criacionistas do design inteligente falem atualmente, cientistas e teólogos refutam sua retórica com um interesse e paixão que não teriam de outra forma. Esta refutação de Luskin, por exemplo, foi criada com a assistência de muitos contribuintes, alguns deles profissionais da área que normalmente não participam de debates criacionistas. No Kansas, e provavelmente em outros estados também, há um interesse na evolução que não estava presente antes da publicação de Ícones. Por qualquer medida, o livro de Wells fez mais para promover a evolução do que para prejudicá-la.
O leitor é novamente lembrado da refutação original de Tamzek a Wells, na qual a flagrante má representação da ciência por Wells é exposta. Como Wells não demonstrou integridade acadêmica, a publicação de seu livro acarretará ou acarretará grandes custos pessoais: Wells não será considerado um investigador ou autor reputado por qualquer grupo, exceto pelos fiéis do design inteligente.
Considerando as bênçãos para a evolução paradoxalmente provindas de Ícones e os custos para a credibilidade de Wells, se a publicação de Ícones como uma "dor no traseiro dos evolucionistas" é uma vitória para Wells, é tanto uma vitória pírrica para ele quanto uma que a ciência e a religião ambas estão felizes em conceder.
Notas finais:
#1: Abell, GO (1983). "As Idades da Terra e do Universo." Cientistas Enfrentam o Criacionismo: Nova York: WW Norton & Company.
#2: Pennock, Robert T. (2000) Torre de Babel: A evidência contra o novo criacionismo. Cambridge: MIT Press. P. 163.
-- Burt Humburg
Nota: Este documento foi editado por várias pessoas, incluindo Nic Tamzek e Mike Hopkins. A introdução original foi escrita por Nic, embora várias outras tenham feito alterações nela.