Resumo: A biologia não pode ser reduzida à física, mesmo que todas as entidades biológicas sejam entidades físicas, e nada mais. A seleção de grupo não é uma teoria evolutiva aceita, mas a ordenação de grupos sim.
A filosofia da ciência, e as críticas à teoria evolutiva em particular, têm sido impulsionadas pela visão de que a física, ou talvez a matemática, é o modelo perfeito de uma disciplina científica moderna. Se não se assemelha à física de alguma forma, então não é ciência.
Como não é surpreendente, muitos biólogos não ficaram satisfeitos com essa visão do que fazem, como se fosse uma espécie de 'coleta de selos' [nota 7]. Ernst Mayr [1970, 1982] atacou essa presunção filosófica, especialmente a ideia de que a biologia é apenas uma forma de física, ou talvez de química. Filósofos também começaram a fazer ataques semelhantes [Hull 1974, ver Sterelny 1995 para uma revisão].
A visão da ciência da filosofia na década de 60 era geralmente reducionista [Nagel 1961]. Isso significava que, em princípio, os objetos e processos de um nível de ciência eram constituídos pelos objetos do nível imediatamente inferior, terminando com a física subatômica. Assim, a biologia se reduzia à química, e a química se reduzia à física. Esse tipo de redução é chamado de redução ontológica. Aqueles que aceitam essa forma de reducionismo são chamados de 'físicistas'.
Outro tipo de redução — frequentemente confundido com a ontológica — é a explicativa ou redução epistêmica. Esta é a visão de que as propriedades de um nível devem ser idealmente explicadas como efeitos de processos no nível imediatamente inferior. Esta visão é amplamente rejeitada por muitos filósofos e biólogos, e defendida por muitos outros [por exemplo, Dennett 1995]. Hull [1974] argumentou que é, em princípio, impossível reduzir, por exemplo, a genética de populações à genética mendeliana, e a genética mendeliana à genética molecular, porque cada nível é o resultado de muitas entidades interagindo no nível inferior, e muitas entidades no nível superior resultam de uma única entidade no nível inferior:
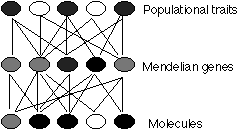
O problema de Hull é às vezes chamado de Problema do Muitos para os Muitos, em homenagem a um antigo problema filosófico, o Problema do Um e dos Muitos. Muitos genes mendelianos são formados por muitas moléculas de DNA, e muitas características populacionais são codificadas por muitos genes mendelianos. Uma simples redução não funcionará. O que Williams [1966] chamou de 'gene evolutivo' é apenas uma unidade de hereditariedade que é 'visível' à seleção, e pode ser uma entidade em qualquer nível - uma molécula, um gene mendeliano ou até mesmo uma característica populacional.
A redução entra no debate evolutivo sob a forma da questão da seleção de grupo. Em 1962, Wynne-Edwards propôs que algumas populações de aves regulam o tamanho do ninho (o número de ovos postos) em tempos difíceis para beneficiar a população como um todo, mesmo que isso fosse prejudicial à 'aptidão darwiniana' dos indivíduos. Williams [1966] respondeu com um argumento de que a seleção de indivíduos não poderia explicar isso e outras formas de suposta seleção de grupo, e que, se a seleção de grupo ocorresse de alguma forma, não seria muito importante.[nota 8] Uma década depois, Dawkins [1976] endureceu essa visão na alegação de que os genes, e apenas os genes, são as 'unidades de seleção', e que todos os efeitos biológicos na evolução são o resultado dessas entidades de 'nível inferior'.
Gene-centrismo não é a visão de que apenas genes existem, ou mesmo que apenas genes têm efeitos, mas que apenas genes são selecionados (isto é, são evolutivamente importantes). A forma como Dawkins colocou, conforme evidenciado pelo título The Selfish Gene, foi mal interpretada para significar que os organismos são irrelevantes. Uma análise mais informada desenvolveu a visão de que, se a evolução por seleção for generalizada, então, usando a própria distinção de Dawkins entre replicadores e veículos (ou o refinamento de Hull, interatores), a seleção pode ocorrer em níveis acima do gene, ou mesmo acima do organismo.
Is refuta noções simplistas de que a evolução é definida como uma mudança na frequência alélica. É isso que Wimsatt chamou de definição de 'contabilidade' da evolução, e é verdadeira até onde vai, mas não é tudo o que há de interessante sobre a evolução. Poucos biólogos são ainda reducionistas simples, embora Williams tenha escrito uma defesa limitada do reducionismo como uma manobra metodológica [1985], na qual ele argumentou que a redução era a 'hipótese nula' e que era oneroso abandoná-la. Dennett [1995] afirmou que a redução ainda não havia falhado, especialmente em explicações evolutivas usando seleção.
Se os níveis evolutivos acima do gene podem ser selecionados, eles estão adaptados? Vários seguiram Wynne-Edwards sobre isso. Versões recentes, no entanto, migraram da noção de que grupos são selecionados, embora, em favor da visão de que eles são classificados, porque a seleção exige que a entidade em questão se replique e o faça de forma diferencial em relação a outros contendores. Isso está bem para genes e, argumentavelmente, para organismos, mas espécies? Até mesmo filos? Grupos não se reproduzem, eles se dividem. Trabalhos recentes de Gould, Eldredge e Vrba [referências em Sterelny 1995] corrigem isso de seleção de espécies seleção para classificação de espécies classificação, o que Vrba chama de 'hipótese do efeito'. Esta é uma visão que agora tem ampla aceitação entre biólogos, incluindo Williams [1992]. Grupos são pensados para sobreviver a eventos de extinção de forma diferencial, com base nas adaptações de seus organismos componentes, então os organismos estão adaptados, não os grupos.