Respuesta a Casey Luskin
por Nic Tamzek et al.[Última actualización: 14 de marzo de 2002]
![]()
Otros enlaces:
|
Introducción
La primera versión del ensayo de Nick Tamzek, "Icono de la Obscuridad", fue un volante bastante largo escrito para ser distribuido en una conferencia de Jonathan Wells en la UCSD el 29 de enero de 2002. El texto de este volante fue publicado en internet poco después, y varias versiones de adiciones y ediciones fueron publicadas sucesivamente hasta que el ensayo fue publicado en talkorigins.org el 25 de febrero de 2002.
Poco después, Casey Luskin del club IDEA de la UCSD, uno de los organizadores de la conferencia del Dr. Wells en la UCSD, publicó una respuesta al volante original (que está alojando aquí) titulado "Íconos que aún permanecen de pie: Jonathan Wells sale limpio, a pesar de la dura crítica". El mismo ensayo ha sido publicado hasta ahora también en el sitio web del centro IDEA de San Diego y en el sitio de ID Access Research Network (ARN).
Aunque el folleto original carecía de gran parte del material que se encuentra en el Archivo TalkOrigins, la extensa respuesta de Luskin intenta abordar muchos de los argumentos clave del FAQ actual, y hasta ahora es la única defensa conocida de Wells. Por lo tanto, una respuesta a Luskin es valiosa. No obstante, se insta al Dr. Wells para que responda por sí mismo.
Esta respuesta ha sido compilada mediante la técnica algo novedosa de que numerosos autores respondan a fragmentos específicos del ensayo de Luskin. Una razón para ello es publicar una versión de este antes del 11 de marzo, cuando Wells y otros hablarán en Ohio. Luskin ha ofrecido una lista tan extensa de afirmaciones creacionistas que una sola persona no puede aclararlas todas en poco tiempo, aunque no sean nuevas. Esto puede hacer que la lectura sea un poco fragmentada, pero permite que diversas personas contribuyan en áreas de conocimiento relevante.
Para mantener las cosas claras, las citas de Luskin estarán en cajas grises como esta. Todas las citas se copian directamente de la versión del ensayo de Luskin que se salvó de la web el 3 de marzo de 2002. Por lo tanto, algunos errores tipográficos en el original se dejarán tal cual, sin [sic], etc. La versión de la defensa de Luskin sobre Wells que se publica en el sitio ARN y en el sitio IDEA cambia constantemente a medida que las personas señalan errores, por lo que esta respuesta necesariamente se refiere a la versión del 3 de marzo. Además, tenga en cuenta que este documento es un trabajo en progreso. Parte de lo que Luskin escribió será abordado en futuras versiones de este documento.
Experimento de Miller-Urey
Abelson continúa diciendo que no hay "evidencia para ... pero mucho en contra" de una atmósfera de amoníaco-metano, señalando que "[u]na cantidad de amoníaco equivalente al nitrógeno atmosférico actual sería destruida [por la radiación ultravioleta] en ~30.000 años."2 Él, y Lasaga et al., encontraron que el gas metano habría dejado una huella de carbono orgánico en las rocas sedimentarias antiguas, quizás incluso debido a una "película de petróleo primordial" de 1 a 10 m de espesor15 (ya que los experimentos de Miller-Urey produjeron por mucho más alquitrán que cualquier otra cosa, y si se produjeron muchos prebióticos, entonces el alquitrán debería haber cubierto la Tierra primitiva), para lo cual no hay evidencia geológica. Schopf señala que el Supergroup de Swazilandia (~3,2 Ga) contiene carbono inorgánico, "indicando que el dióxido de carbono, y no el metano, pudo haber sido la forma dominante de carbono atmosférico en este punto en el tiempo"16. Esto desafía la importancia de las fuentes hidrotermales submarinas como contribuidoras de metano, lo que Tamzek dice que Wells "excluye" incorrectamente.
Como esto ocurre alrededor de 300 millones de años después de los primeros fósiles bacterianos conocidos, y mucho después del período de tiempo del origen de la vida (4,2-3,8 Bya), ¿alguien se sorprende? Además, en el modelo de Kastings, el metano es una forma menor (alrededor de 20 ppm) y no dominante en la atmósfera, por lo que esto es doblemente irrelevante para el modelo de chimeneas hidrotermales.
-- Ian Musgrave
El árbol de la vida de Darwin
El reloj de Darwin está sonando
Tamzek señala que la hipótesis del reloj molecular está en gran disputa, lo cual es más pronunciado cuando el reloj molecular difiere marcadamente del registro fósil (especialmente con respecto a los mamíferos36, los pájaros36 y los grandes grupos animales36, 37). Los caracoles cone79, que Tamzek alude como un ejemplo de evolución rápida, proporcionan poco material para refutar las afirmaciones creacionistas contra el origen natural de la información genética, ya que simplemente muestra que hubo una fuerte presión de selección sobre el mrRna de los caracoles cone. Si la evolución ocurrió, no se crearon nuevas funciones aparte de unos pocos venenos destructivos diferentes, y estos venenos mismos han tenido entre 20-50 Ma para desarrollarse en sus formas bastante similares. El papel admite en realidad que su ejemplo más llamativo de cambio rápido alegado, al que llama una "hipermutación focal"79, es "mecánicamente inexplicado", haciendo que las explicaciones naturalistas, en este punto, sean impotentes para explicar las transformaciones genéticas alegadas.
Cuando Luskin se pone a abordar la duplicación génica como un ejemplo de la evolución aumentando la información, tiene algunas cosas extrañas que decir.
Este artículo, como muchos otros en la literatura de la biología evolutiva, basa la mayor parte de su evolución genética de la bioquímica en el mecanismo de la duplicación génica. Sin embargo, al intentar evolucionar algo, la explicación de la duplicación génica no ayuda mucho con el problema, porque una vez que duplicas un gen, tienes un nuevo pedazo de información genética con el que jugar, pero ¿qué utilidad tiene eso para ti? Si los sistemas complejos necesitan partes específicas, ¿qué tipo de evidencia hay de que estos genes duplicados serán las partes que necesitas?
Por supuesto, las duplicaciones génicas ocurren en organismos ya vivos que no "necesitan" nada. Sin embargo, si una duplicación produce una función adicional útil o amplía una función existente, entonces la selección natural ayudará a que se propague. Esta es una de las formas más frecuentes en que se añade nueva información funcional a los genomas, y los nuevos sistemas complejos son el resultado eventual de nuevos genes trabajando juntos.
Luskin basa la mayor parte de su rechazo en un artículo de Lynch y Conery [1], que afirma que la tasa de duplicación génica es demasiado baja, pero su uso del artículo presenta varios problemas graves:
1. Lynch y Conery, al utilizar genomas completos, han intentado derivar empíricamente la tasa de duplicaciones de genes individuales, y lo han hecho excluyendo familias de genes múltiples. Su estudio no abordó la duplicación del genoma completo, la duplicación parcial o completa de cromosomas, ni otros tipos de duplicación segmental. Luskin descarta la duplicación como un medio para generar novedad biológica, pero falla al abordar los mecanismos más potentes para la adición de material genético, los cuales son capaces de añadir cientos o miles de genes a la vez.
2. Contrario a la afirmación de Luskin de que la tasa de duplicación reportada es "bastante rara", en realidad se considera bastante alta. Lynch y Conery afirman que "el 50% de todos los genes en un genoma se espera que se dupliquen y aumenten a alta frecuencia al menos una vez en escalas de tiempo de 35 a 350 millones de años. Por lo tanto, incluso en ausencia de la amplificación directa de genomas enteros (poliploidización), la duplicación génica tiene el potencial de generar un sustrato molecular sustancial para el origen de novedades evolutivas."
3. Las metodologías y conclusiones de Lynch y Conery fueron ambas fuertemente criticadas en un artículo de "comentarios técnicos" de Science por dos conjuntos separados de científicos [2], [3] que son expertos muy reconocidos. Entre otras cosas, se cuestionó la conclusión sobre la vida media corta de los duplicados génicos y la tasa de duplicación, y aunque Lynch y Conery defienden su postura, admiten que varias suposiciones sesgan sus resultados y revisan algunas de sus estimaciones. Por lo tanto, los resultados del estudio son al menos altamente cuestionables. Luskin debería haber mencionado esta controversia, o haber omitido el estudio por completo.
Incluso sin estos problemas, Luskin sigue sin abordar adecuadamente el problema de la duplicación al basarse en este estudio. La tasa que presentan Lynch y Conery, incluso si es correcta, es una tasa promedio de duplicación. Pero diversos genes pueden desviarse considerablemente de la norma. Además, es bien sabido que las tasas de duplicación génica no son uniformes. Por ejemplo, las duplicaciones subsiguientes son mucho más probables si ha ocurrido una duplicación previamente, porque las posibilidades de un entrecruzamiento desigual aumentan considerablemente [4]. Esto permite la rápida expansión de familias génicas, lo cual parece haber ocurrido en el caso de los venenos del caracol cono. Lynch y Conery dejaron fuera de su estudio las familias génicas.
Lynch y Conery encontraron que el promedio de duplicaciones génicas ocurre aproximadamente una vez cada 100 millones de años108; eso es bastante raro. Si las caracolas cónidas tienen un tiempo de generación de 1 año, y el gen que necesitas se duplica una vez cada 100.000.000 de generaciones, eso no te ofrece una muy buena oportunidad de obtenerlo cuando lo necesitas. Además, se ha encontrado que "la gran mayoría de las duplicaciones génicas se silencian dentro de unos pocos millones de años, y los pocos supervivientes posteriormente experimentan una fuerte selección purificadora"108. Otro estudio mostró que no son muy libres de mutar en absoluto, que existe una fuerte presión selectiva sobre ellas107. Esto apoya la afirmación de Conery y Lynch de que los mecanismos reales mediante los cuales la duplicación génica contribuye a la evolución no están muy bien comprendidos:
"Sin embargo, no está claro cómo los genes duplicados navegan exitosamente una trayectoria evolutiva desde un estado inicial de redundancia completa, en el que una copia probablemente sea prescindible, hasta una situación estable en la que ambas copias son mantenidas por selección natural. Tampoco está claro con qué frecuencia ocurren estos eventos."108
Luskin se basa en Lynch y Conery para afirmar que los nuevos duplicados no duran mucho porque son silenciados rápidamente (es decir, en millones de años) por mutación. (Ironicamente, los que se ven silenciados se convierten en pseudogenes, los cuales ofrecen evidencia poderosa de la descendencia común. Véase "Errores plagiarizados y genética molecular", http://www.talkorigins.org/faqs/molgen/). Pero aparte de los problemas mencionados anteriormente con ese estudio, Luskin simplemente ignora estudios recientes que muestran que los duplicados se conservan mucho más a menudo de lo que se pensaba anteriormente [5], [6], incluyendo estudios realizados por Lynch mismo [7], [8]. Además de evolucionar funciones completamente nuevas, las duplicaciones génicas pueden repartir funciones antiguas que el gen ancestral llevaba a cabo solo débilmente. Este modelo está respaldado por el hecho de que muchas enzimas son "promiscuas" en cuanto al número de actividades débiles [9]. Esto permite que las funciones repartidas sean más activas y estén reguladas de manera diferente, a veces siendo expresadas en diferentes tejidos [10]. De esta manera, puede evolucionar una mayor complejidad incluso sin añadir funciones de novo, y los duplicados se conservan con mayor frecuencia. Luskin concluye:
Lo que queda claro es que la explicación de la duplicación génica sigue dejando los detalles al azar, y esta vía definitivamente no ha sido verificada experimentalmente. Todo lo que Espiritu et al. han encontrado son homologías de proteínas, y luego han inferido una vía ancestral vaga de creación génica. Esta explicación para el origen de la verdadera novedad evolutiva carece de un mecanismo confiable y es poco mejor que una especulación sin fundamento.
Pero es claro que es Luskin quien está haciendo gestos vacíos. Se basa en un estudio cuestionable que interpreta mal mientras ignora una abundancia de datos que contradicen su tesis. Contrario a sus afirmaciones, los mecanismos para la duplicación génica, que incluyen el entrecruzamiento desigual y la inserción de retrogenes, son razonablemente bien comprendidos [4] y son seguidos por cualquier número de posibilidades selectivas (véase cualquiera de las referencias anteriores o siguientes para información extensa, o cualquiera de las decenas de otras). Además, la duplicación génica adaptativa ha sido observada en el laboratorio [11], [12]. Y observada entre especies estrechamente relacionadas [13] (incluyendo caracoles cone [14]) que incluso los creacionistas de la Tierra joven consideran relacionadas por descendencia común. Ciertamente, hay más cosas por aprender sobre la duplicación génica (lo cual es una razón por la cual los venenos de caracoles cone son tan intensamente estudiados), pero si ocurre o no, o su importancia en la evolución, no está en duda. Simplemente es incorrecto afirmar que la duplicación génica no es un mecanismo confiable para generar novedad biológica.
REFERENCIAS
1. Lynch, M. y J.S. Conery, El destino evolutivo y las consecuencias de los genes duplicados. Science, 2000. 290(5494): p. 1151-5.
2. Long, M. y K. Thornton, Duplicación génica y evolución. Science, 2001. 293(5535): p. 1551.
3. Zhang, L., B.S. Gaut, y T.J. Vision, Duplicación génica y evolución. Science, 2001. 293(5535): p. 1551.
4. Patthy, L., Evolución de proteínas. 1ª ed. 1999: Blackwell Science Inc. 91-103.
5. Massingham, T., L.J. Davies, y P. Lio, Análisis de la función génica tras la duplicación. Bioessays, 2001. 23(10): p. 873-6.
6. Dermitzakis, E.T. y A.G. Clark, Selección diferencial tras la duplicación en genes del desarrollo mamífero. Mol Biol Evol, 2001. 18(4): p. 557-62.
7. Force, A., et al., Preservación de genes duplicados mediante mutaciones complementarias y degenerativas. Genética, 1999. 151(4): p. 1531-45.
8. Lynch, M. y A. Force, La probabilidad de preservación de genes duplicados mediante subfuncionalización. Genética, 2000. 154(1): p. 459-73.
9. James, L.C. y D.S. Tawfik, Reactividades poliméricas catalíticas y de unión compartidas por dos proteínas no relacionadas: El potencial papel del promiscuidad en la evolución enzimática. Protein Sci, 2001. 10(12): p. 2600-7.
10. Serluca, F.C., et al., La partición de la expresión tisular acompaña múltiples duplicaciones del gen de la subunidad alfa de la Na+/K+ ATPasa. Genome Res, 2001. 11(10): p. 1625-31.
11. Jelesko, J.G., et al., Recombinación desigual germinal rara que conduce a la formación de genes recombinantes y duplicación génica en Arabidopsis thaliana. Proc Natl Acad Sci U S A, 1999. 96(18): p. 10302-7.
12. Brown, C.J., K.M. Todd, y R.F. Rosenzweig, Duplicaciones múltiples de genes de transporte de hexosas de levadura en respuesta a la selección en un ambiente limitado en glucosa. Mol Biol Evol, 1998. 15(8): p. 931-42.
13. Cirera, S. y M. Aguade, Evolución molecular de una duplicación: la región del gen del péptido sexual (Acp70A) de Drosophila subobscura y Drosophila madeirensis. Mol Biol Evol, 1998. 15(8): p. 988-96.
14. Espiritu, D.J., et al., Caracoles cónicos venenosos: filogenia molecular y la generación de diversidad de toxinas. Toxicon, 2001. 39(12): p. 1899-916.
-- Steve Reuland
¡Salva los árboles!
Tamzek sugiere que el árbol de la vida de Darwin parece simple y "arbolado" aparte del 'bosque molecular' en la raíz, sin embargo esto no es así. Está bien reconocido en sistemática que muy a menudo los árboles filogenéticos basados en una sola secuencia de gen o proteína, darán como resultado un árbol, mientras que un árbol basado en alguna otra biomolécula parecerá bastante diferente. Y aún más frecuentemente, los árboles basados en biomoléculas entran en conflicto con los árboles creados a partir de características macromorfológicas o el registro fósil (y los árboles basados en diferentes macromorfologías a menudo difieren entre sí bastante). Tamzek afirma que el árbol de los Eucariotas está bien resuelto a través de Baldauf et al. (2000)99, sin embargo este artículo escapa a los problemas típicos de árboles en conflicto porque crea un árbol utilizando un conjunto de datos masivo de muchas secuencias de proteínas para ocultar estadísticamente las diferencias entre los árboles basados en proteínas individuales. Al crear un árbol "flaco" a través de muchos genes, la ancestría común no puede ser fuertemente verificada a través de líneas de evidencia genética independientes (reconociendo que esto es una buena técnica para crear una filogenia si uno ya asume que la ancestría común es verdadera, sin embargo, como una prueba de la ancestría común, muestra que la evidencia molecular proporciona poco apoyo). De hecho, se encontró que las filogenias de un solo gen solo "apoyan subconjuntos del árbol de proteínas combinado". En otras palabras, los genes diferentes, cuando se toman independientemente, no convergen para formar un árbol bonito y ordenado.
La afirmación de Luskin de que Baldauf et al. combinaron los datos para oscurecer el conflicto es infundada. Los autores reconocieron explícitamente el potencial de conflicto entre los conjuntos de datos, y para abordar este problema, analizaron las proteínas por separado antes de combinarlas. Aquí está lo que Baldauf et al. escribieron:
Los análisis de proteínas individuales muestran que no se apoyan agrupaciones contradictorias con >65% aaBP y la mayoría con mucho menos (Fig. 3, columnas 1 a 4). Por lo tanto, no existen conflictos fuertemente apoyados entre estos datos. Los análisis por pares también muestran considerable evidencia de cooperación filogenética entre todas las particiones para la mayoría de las preguntas (Fig. 3, columnas 5 a 10); es decir, la mayoría de las combinaciones por pares tienden a apoyar las mismas conclusiones. Esto es más evidencia de historias subyacentes comunes entre todas las particiones. [Énfasis añadido.]
Baldauf et al. no combinaron los datos para ocultar un conflicto porque no encontraron un conflicto significativo. Los clados que parecían entrar en conflicto no tenían soporte estadístico y probablemente fueron el resultado de datos insuficientes.
- Alex Wild
Tamzek afirma que los resultados de este artículo indican que la filogenia de los Eucariotas está avanzando "muy bien", y que este documento proporciona una filogenia de los Eucariotas "perfectamente tradicional y en forma de árbol". Como se dijo anteriormente, la limpieza de este árbol es puramente el resultado de las técnicas estadísticas y el conjunto de datos combinado utilizado en el artículo.
Parece que debemos añadir la estadística a la lista de disciplinas que deben estar defectuosas si los argumentos del Diseño Inteligente son ciertos. La principal "técnica estadística" utilizada para producir la "limpieza" del árbol de Baldauf et al. es el uso de una muestra más grande de datos de caracteres. Si extraer de una muestra más grande es una técnica estadística deshonesta, los cerdos vuelan.
Según la lógica de Luskin, deberíamos rechazar el resultado de una encuesta de opinión de 100 personas en favor del resultado de una encuesta de opinión de 10 personas, porque la primera utilizó una "técnica estadística" que produjo un resultado diferente. Luskin nos pide que pesemos un resultado débilmente apoyado basado en una muestra pequeña igual que un estudio mucho más robusto basado en una muestra más grande.
¿Por qué hace esto? No lo sé. Supongo que el error de muestreo produce suficientes resultados extraños para que los defensores del diseño inteligente encuentren algunos que usar con fines retóricos, especialmente si evitan los problemas del tamaño de la muestra y el poder estadístico. Sin embargo, una vez que comencemos a evaluar estudios no por los méritos de su metodología sino por lo bien que podamos usarlos en nuestra retórica, dejamos de ser buenos académicos y comenzamos a ser propagandistas.
-- Alex Wild
De todos modos, Baldauf et al. encontraron de hecho que su árbol presenta "diferencias notables con la filogenia del ARNr SSU"99, y señalaron que estudios anteriores también han encontrado árboles contradictorios.
Veamos el resumen del artículo de Baldauf et al., la fuente de la cita:
La comprensión actual de la sistemática de alto orden de los eucariotas se basa en gran medida en análisis del ARN de la subunidad ribosomal pequeña (SSU rRNA). La verificación independiente de estos resultados sigue siendo limitada. Hemos combinado las secuencias de cuatro de las proteínas más ampliamente muestreadas taxonómicamente disponibles para crear un conjunto de datos aproximadamente paralelo al del SSU rRNA. El árbol filogenético resultante muestra una serie de diferencias notables con respecto a la filogenia del SSU rRNA, incluyendo un fuerte apoyo para la mayoría de los grupos principales y varias supergrupos principales. [Énfasis añadido.]
Observe en particular la frase que Luskin omitió:
...incluyendo un fuerte apoyo para la mayoría de los grupos principales y varios supergrupos principales
El texto omitido cambia significativamente el significado de la oración. Luskin estaba haciendo el caso de que el ARNr SSU proporciona un árbol diferente al producido por los 4 proteínas utilizadas por Baldauf et al., y en particular estaba argumentando que los dos conjuntos de datos producen árboles conflictivos. Sin embargo, en contexto es claro que las diferencias entre el ARNr SSU y los árboles de Baldauf et al. no están principalmente en el orden de ramificación, sino en el nivel de soporte para los agrupamientos principales. Una lectura adicional de Baldauf et al. confirma esto. La diferencia entre un soporte fuerte y un soporte débil para un agrupamiento (o de un soporte fuerte para un agrupamiento versus un soporte débil para un agrupamiento diferente) es sin duda alguna diferente de la diferencia entre dos agrupamientos fuertemente soportados pero conflictivos. El argumento de Luskin se basa en confundir los dos.
Fuente: Baldauf SL, Roger AJ, Wenk-Siefert I, Doolittle WF. "Una filogenia a nivel de reino de eucariotas basada en datos combinados de proteínas." Science 290(5493):972-7 (3 de noviembre de 2000).
-- Alex Wild
Embriones de Haeckel:
Developmental biologist PZ Myers was asked via email to comment on what Luskin wrote:
La Figura de la Hora Embriológica
Tamzek concede graciosamente que los dibujos de Haeckel no pertenecen en los libros de texto (afirmando esencialmente el ícono), sin embargo, cita a Raff (2001)30 para implicar que la embriología aún proporciona evidencia de una historia evolutiva compartida entre las clases de vertebrados. Raff implica que Wells niega la "fase filotípica" y el "compartir de elementos estructurales principales y sus elementos topológicos" entre los embriones de vertebrados, sin embargo Wells reconoce fácilmente que las similitudes embrionarias de la "fase filotípica" sí existen diciendo: "aquí [en la fase filotípica] las diversas clases exhiben por primera vez las características comunes a todos los vertebrados" (Íconos, pág. 98). El punto de Wells, aquí, es que las rutas que toman los vertebrados en desarrollo para llegar a la "fase filotípica" son muy diferentes, formando un patrón de desarrollo que Raff mismo dice tiene forma de "hora"32. Wells señala que este patrón de "hora" no se predice si todos los vertebrados comparten un ancestro común.
Esto es correcto. Un patrón específico de desarrollo no es una predicción evolutiva; la ampolla del desarrollo es una observación. Observamos un periodo filotípico una y otra vez en diferentes especies, lo que plantea una pregunta interesante: ¿por qué hacen eso los animales? La explicación evolutiva es que existe una base morfológica (el plan corporal) que se establece en un periodo temprano del desarrollo, la cual ha sido refractaria al cambio. Las elaboraciones posteriores del plan corporal se basan en los patrones establecidos en la etapa filotípica, por lo que esa etapa, definida de manera amplia, persiste.
Además, podemos hacer predicciones basadas en hipótesis sobre cómo se establecen los planes corporales. Sabemos que los genes Hox son centrales en este proceso. Una predicción que se puede hacer es que los organismos con diferentes planes corporales deberían tener cambios correlacionados en la forma en que se expresan sus genes Hox, o en la forma en que los genes Hox regulan genes más aguas abajo. Estas predicciones han sido probadas y actualmente se están probando en laboratorios de todo el mundo.
El campo del creacionismo/diseño inteligente carece de cualquier explicación para las características comunes que observamos en organismos ampliamente dispares. Si sus 'teorías' tuvieran algún poder, deberían ser capaces de ayudarnos a comprender estas observaciones. No lo hacen.
Myers:
[...]
Richardson dice que esto no es un problema para la evolución darwiniana porque "la mezcla de similitudes y diferencias entre embriones de vertebrados refleja el cambio evolutivo en los mecanismos del desarrollo heredados de un ancestro común". Sin embargo, al examinar los datos que Wells presenta en su capítulo sobre la homología en las extremidades de los vertebrados, encontramos que los mecanismos que controlan el desarrollo a menudo no se predicen en absoluto por la filogenia. Wells encuentra que el gen Distal-less controla el desarrollo de las apéndices en el ratón, el gusano espinoso, la mariposa, el erizo de mar y el gusano terciopelo, a pesar de que tales apéndices no se derivan a través de una descendencia común. No hay razón por la cual la descendencia común debería predecir que el mismo gen produzca "patas" en tantos tipos muy diferentes de organismos, cuando su supuesto último ancestro común no tenía patas.
Esto es parcialmente cierto. Es correcto que no predeciríamos que las patas en artrópodos y vertebrados necesariamente tendrían la misma causa molecular, porque las patas en esos dos linajes no son homólogas: surgieron independientemente. Sin embargo, lo que tenemos aquí nuevamente es una observación. Las patas de vertebrados y artrópodos utilizan todas el mismo gen, distal-less, para iniciar algunas de las primeras etapas en el desarrollo de las extremidades. ¡Ese es un resultado bastante sorprendente que favorece una explicación evolutiva! Los evolucionistas siempre han predicho algún nivel no especificado de similitud entre diferentes linajes, porque surgieron de un ancestro común, pero no podíamos saber cuán similares serían hasta que los miráramos. Ahora estamos en la curiosa posición en la que los creacionistas intentan argumentar que, porque las similitudes entre los organismos son más fuertes de lo que habíamos anticipado, eso es evidencia en contra de la idea de la descendencia común!
Archaeopteryx: El eslabón perdido:
Vertebrate paleontologist Thomas R. Holtz, Jr. who showed just how inaccurate Luskin is. Holtz wrote in an email:
Fundamentado.
Tamzek afirma que los terópodos fósiles Dromaeosauria (Protoarchaeopteryx y Caudipteryx) proporcionan evidencia inequívoca de que las aves evolucionaron a partir de dinosaurios.
Ni Protarchaeopteryx (nota la ortografía) ni Caudipteryx son dromeosaurios. Son un maniraptorano generalizado y un oviraptorosaurio, respectivamente.
En el resto de esta sección, las réplicas a Luskin entre comillas provienen del correo electrónico de Holtz. Éste proporcionó el siguiente cladograma:

"En este diagrama, los siguientes grupos tienen plumas confirmadas/análogos de plumas:
Avialae (aves) (eh...)
Dromaeosauridae (Microraptor,
Sinornithosaurus, y la nueva forma mencionada en
Nature esta semana)
Alvarezsauridae (Shuvuuia)
Oviraptorosauria (Caudipteryx)
Therizinosauroidea (Beipiaosaurus)
Compsoganthidae (Sinosauropteryx)
Observe que, con excepción de Shuvuuia y las aves, todos los
descubrimientos provienen de una sola Formación, el Yixian,
conocida por su espectacular preservación. Observe también que
no tenemos impresiones de escamas o cualquier otra impresión
no plumaria para otras especies en estos grupos particulares,
por lo que la explicación más simple es que su ancestro común
más reciente tenía plumas simples."
"The dinosaurs from Tyrannosauroidea up to birds on this cladogram are noted by a reduction in the number of tail vertebrae; this is particularly true of birds and their closest relatives."
Sin embargo, el escéptico debe hacerse la pregunta: ¿es esto realmente así? Estos son fósiles interesantes, pero no proporcionan suficiente evidencia para que la hipótesis del dinosaurio-ave despegue. De hecho, las colas pesadas y los grandes cuerpos de los dinosaurios terópodos son exactamente lo opuesto al tipo de organismo que esperaríamos que evolucionara hacia un volador92.
"These guys are mixed up; the description they refer to sounds like Sinosauropteryx (a compsognathid). No one has questioned the feathers on Protarchaeopteryx; the only question was whether or not it was a bird."
Según el informe en Nature38, Protoarchaeopteryx era un dinosaurio con plumas similares a "pelo" en su cuerpo, sin embargo un paleontólogo llamó a ese pelo "dino-pelo, [que] realmente podría no tener nada que ver con el origen de las plumas."91
De todos modos, las plumas de contorno no tienen función para el vuelo en las aves. Las únicas "plumas de contorno, con barbas y simétricas"38 en Protoarchaeopteryx aparecen en su cola: no hay signos de plumas para el vuelo. Caudipteryx tiene algunas plumas de contorno y barbas en un dedo de la mano, pero su brazo es mucho más corto que el de un ave39: demasiado corto para el vuelo. Dado que estos organismos tienen algunas estructuras corporales bien adaptadas al vuelo, pero claramente no volaron, estos (y otros) fósiles han llevado a los evolucionistas a creer que algunas de las estructuras complejas principales especificadas para el vuelo —plumas, alas y huesos óseos— se originaron con un propósito distinto al vuelo40.
Ninguno de estos dos tiene 'alas'; son solo brazos. Los de Caudipteryx, de hecho, son realmente cortos en comparación con otros oviraptorosaurios.
"'huesos óseos' = huesos hechos de huesos. Dado que tú y yo tenemos huesos óseos, al igual que los gatos, los perros, los lagartos, las ranas y las truchas, esto apenas es una característica de vuelo!!"
No hay explicaciones elegantes aquí, sino pensamientos deseosos que intentan forzar los datos a un modelo de evolución que falla al explicar el origen del vuelo.
"En realidad, lo que hay es la descripción del material a mano. Fósiles primero, chicos; hipótesis segundo."
"Nope. These have been shown to be structurally disctinct from the true feathers of birds and other dinosaurs; see recent papers by Reisz and Sues."
Reptiles de plumas, no se reúnen en bandadas
De hecho, las plumas también se han encontrado en un reptil similar a un lagarto no dinosaurio90,
sin embargo, los defensores de la hipótesis del dinosaurio-ave piensan que este fósil emplumado está completamente desconectado del origen de las aves y de los supuestos dinosaurios emplumados. ¿Entonces, por qué debería la supuesta aparición de plumas en estos dromaeosaurios post-aves convertirse en la prueba definitiva de la ascendencia dinosauriana de las aves? ¿Quizás estos "dinos emplumados" son simplemente lo que sus escamados
"Longisquama NO es un escamado!!! (Escamado = lagartos y serpientes). Tampoco es un arcosaurio verdadero, como tradicionalmente se ha considerado.
"Además, la evidencia principal del origen dinosauriano de las aves es esquelética, no integumentaria. Consulte mi sitio web, entre muchas otras referencias."
"Identical."
sus contrapartes son: quimeras de reptiles no relacionadas con las aves.
Para arrojar algunas otras piedras en la hipótesis dino-ave, aunque tanto los dinosaurios terópodos como las aves caminan sobre 2 patas y tienen algunas similitudes esqueléticas, diferencias como la configuración de los dígitos,
"identical (and in fact the same thing: pubis bone being part of the pelvis!) in basal dromaeosaurids, basal troodontids, and basal birds."
hueso del pubis, forma del pelvis,
"Teeth of primitive birds, primitive troodontids, and primitive dromaeosaurids all very similar."
dientes,
"Fair to say that it is distinct from all LIVING reptiles."
y la disposición de los órganos internos (el sistema respiratorio aviar es único y muy diferente al de los reptiles)
And rejected...
han sido todas planteadas
como diferencias que desafían la hipótesis dino-ave43, 92. Wells también señala que los métodos cladísticos que han establecido esta supuesta relación ignoran el registro fósil,
"Necio. ¿Cómo podría un análisis cladístico de formas conocidas únicamente a partir de fósiles ignorar el registro fósil?
"(Sospecho que se refería al 'registro estratigráfico', pero eso es otro asunto)"
"Guilty. :-) "
asuma un historial evolutivo,
y ignore los problemas importantes con las implicaciones de que los dinosaurios evolucionaron para volar desde el suelo.
"Los análisis filogenéticos son hipótesis de relaciones; son el marco mediante el cual otras investigaciones (como el origen del vuelo o similares) podrían interpretarse, pero los análisis en sí mismos son SOLO las interrelaciones."
El texto completo del correo electrónico de Holtz está aquí.
Polillas del carbón y los pinzones de Darwin:
Ya basta con mi retórica creacionista: ¿tiene razón Wells?
En lo que respecta a las polillas, Tamzek cita a Majerus, quien, en respuesta a Wells, admite que la hipótesis del melanismo está "descreditada", lo cual es efectivamente lo que se enseña en los libros de texto, validando así la crítica general de Wells. En esta respuesta de 2000 a Wells, citada por Tamzek, Majerus afirma que la depredación diferencial por parte de las aves sigue siendo responsable del cambio en la coloración de las polillas. Sin embargo, el mismo Majerus escribió en 1987: "parece seguro que la mayoría de los B. betularia descansan donde están ocultos.... [y] que las áreas expuestas de los troncos de los árboles no son un sitio de descanso importante para ninguna forma de B. betularia"87, lo que excluiría la posibilidad de que la depredación por parte de las aves sea un factor importante en su selección. Majerus, un experto, parece estar contradiciéndose aquí o retractando sus afirmaciones.
En la correspondencia citada, Majerus no dice que la idea de que las aves que se alimentan de polillas camufladas de manera diferencial resulte en melanismo industrial fue "descreditada", sino que afirma explícitamente que existe evidencia sustancial a su favor. Él sí dijo que la presentación en los libros de texto es excesivamente simplificada, lo cual no valida la crítica de Wells.
Si el autor hubiera leído realmente la referencia 87 (Howlett R.J., Majerus M., Biological Journal of the Linnean Society 30: 31-44 (1987)), habría notado que Majerus y Howlett no discutieron que la depredación por aves fuera un factor importante en la selección en ese artículo. Su preocupación fue obtener valores cuantitativos precisos para la selección mediante la depredación por aves. El autor también debería haber notado que Howlett y Majerus reportaron los resultados de un experimento comparando la presión de selección en lugares expuestos y no expuestos. No hubo diferencia significativa en la proporción de polillas oscuras y claras capturadas por aves entre lugares expuestos y no expuestos, por lo tanto, la depredación por aves es un factor importante en ambos lugares. La postura de Majerus en 1987 fue la misma que en 2000.
-- Ian Musgrave
Independientemente de lo que hagan realmente las polillas, Wells no estaba injustificado al confiar en Majerus, un experto (y otros88), cuando Majerus sugirió que las aves no son un factor en la selección de las polillas87. Majerus y otros pueden discrepar con Wells, sin embargo, Wells mismo no está completamente ignorante sobre el tema de las polillas, ya que publicó un artículo de opinión sobre el tema titulado, "Second Thoughts about Peppered Moths"88 en la revista revisada por pares, The Scientist.
La afirmación de revisión por pares es una declaración falsa. The Scientist se describe como "La revista de noticias para las ciencias de la vida" (http://www.the-scientist.com/). En una breve búsqueda del sitio web, no puedo encontrar nada que mencione la revisión por pares o la falta de la misma, ni ninguna guía de envío. Esto no es una buena señal. En cualquier caso, el artículo de Wells estaba en la sección de "Opinión", como puedes ver aquí mismo.
También he enviado un correo electrónico a The Scientist para solicitar aclaraciones sobre la revisión por pares. El editor, Alexander M. Grimwade Ph.D., respondió:
1. THE SCIENTIST no es una revista revisada por pares.
2. El artículo escrito por Wells fue publicado en nuestra sección de Opinión, donde permitimos a los científicos "expresarse" con una edición o revisión del contenido mínima. Para citar nuestro encabezado:
"THE SCIENTIST sirve a sus lectores de muchas maneras, pero uno de sus aspectos más valorados es su compromiso con la discusión abierta de temas controvertidos. Si bien los lectores a menudo nos elogian por este compromiso, también deberían reconocer que todos los artículos publicados en THE SCIENTIST reflejan las opiniones de sus autores y no las opiniones oficiales de la publicación, su personal editorial o su propiedad."
El hecho de que The Scientist no haya comprendido completamente la situación de las polillas al aceptar el artículo de Wells se indica mediante su viñeta. ¿Quién iría a cazar polillas con bata de laboratorio y lupa?
Además, la versión original de la respuesta de Luskin decía que el artículo de Wells en The Scientist "contenía más de 45 referencias" —en realidad tenía 17. La versión ampliada del ensayo de la polilla de Wells está disponible en ARN, que es probablemente a lo que se refería Luskin, aunque un conteo cuidadoso de la lista de referencias revela 44 referencias. En cualquier caso, el número de referencias es irrelevante si se las utiliza de manera injusta, como muestra la versión ampliada de la sección sobre la polilla del tabaco de mi artículo.
Si, sin embargo, eres un verdadero glotón de detalles bien referenciados sobre polillas del hollín y Wells, entonces la persona a consultar es Ian Musgrave. Algunos de sus comentarios están en línea aquí.
-- Nic Tamzek
Majerus nunca implicó que la depredación por aves no fuera un factor importante en la selección. De hecho, él establece explícitamente que lo es, y enfatiza que su preocupación es obtener una estimación cuantitativa precisa de la presión selectiva debida a la depredación por aves. Que Wells fuera capaz de publicarse en "The Scientist" no dice nada sobre su conocimiento de las polillas. "The Scientist" es no una publicación revisada por pares, sino una publicación dedicada a artículos de interés general y noticias. Más que un examen superficial del artículo de Howlett y Majerus habría mostrado que Majerus no estaba disputando que la depredación por aves fuera un factor importante (aunque no necesariamente exclusivo) en el melanismo industrial. No es que esto sea sorprendente, ni siquiera Kettlewell pensaba que la depredación fuera el único factor involucrado (Kettlewell, HBD. (1961) Ann. Rev. Ent., 6: 245-262.).
-- Ian Musgrave
No quiero añadir más, pero este punto no puede ser enfatizado lo suficiente. Majerus ha repetido y claramente ha expresado su opinión de que las historias de los libros de texto son excesivamente simplificadas, pero que el punto fundamental, que es que la depredación de mariposas por parte de aves es la fuerza selectiva principal que causa el cambio en el color de las mariposas, sigue siendo una buena conclusión bien respaldada por los datos.
Aquí están las conclusiones de Majerus sobre el caso de la polilla del roble, al final de los dos capítulos (casi sesenta páginas) de su libro Melanismo industrial que dedica precisamente a este tema:
El caso de la polilla del abedul es sin duda más complejo y fascinante de lo que la mayoría de los libros de texto de biología tienen espacio para tratar. Incluso aquí, he tenido que ser selectivo sobre qué conjuntos de datos incluir, dada la enorme cantidad de material sobre este caso. He tendido a concentrarme en trabajos realizados en el noroeste industrial de Inglaterra, porque es el trabajo más extenso, y en East Anglia, donde he trabajado yo mismo. Mi objetivo en este y el capítulo anterior ha sido proporcionar suficiente información para que el lector pueda evaluar por sí mismo qué partes de la historia están completas y dónde se necesita más trabajo (véase el Capítulo 10). Mi visión del ascenso y caída de la polilla del abedul melánica es que la depredación diferencial por parte de aves en regiones más o menos contaminadas, junto con la migración, son principalmente responsables, casi a la exclusión de otros factores. (Majerus, 1998, Industrial Melanism, p. 155, énfasis añadido)
-- Nic Tamzek
Del Simio al Humano: El Ícono Supremo:
Anthropologist Jeffrey K. McKee, author of La Cadena Rellena: Azar, Coincidencia y Caos en la Evolución Humana, provided a reply to this via email:
El paleontólogo Stephen Jay Gould probablemente estaría de acuerdo con Wells, ya que una vez escribió: "[l]a mayoría de los fósiles de homínidos, aunque sirven como base para especulaciones interminables y elaboradas historias, son fragmentos de mandíbulas y trozos de cráneos."48 Roger Lewin también estaría de acuerdo cuando escribió: "el problema principal ha sido el lamentablemente pequeño número de fósiles de homínidos sobre los cuales los prehistoriadores podrían ejercer sus talentos interpretativos"75. Wells no es en absoluto deshonesto al afirmar que la escasez de fósiles útiles para investigar la presunta ascendencia homínida de los humanos causa subjetividad entre los investigadores. Las declaraciones de Wells están en línea con lo que Gould y Lewin han dicho, y no son negadas por un nivel reducido de relevancia encontrado en la cita de Wells a Henry Gee.
Si se intenta establecer evidencia de que los humanos evolucionaron, la verdad es que los paleoantropólogos tienen un exceso de riquezas. Hay miles de fósiles con los que documentar las tendencias principales de los últimos 4 millones de años, comenzando con las primeras criaturas bípedas (Australopithecus) que conservaron las extremidades superiores para escalar y solo tenían un escaso aumento del tamaño del cerebro en comparación con los simios vivos. Los primeros Homo (es decir, Homo habilis) mostraron aumentos adicionales en el tamaño relativo del cerebro, y aunque las extremidades cambiaron sutilmente, sus proporciones de extremidades permanecieron similares a las de los simios. Homo erectus/ergaster llevó las tendencias más lejos, con homínidos que alcanzaron un tamaño corporal moderno y proporciones corporales generales, pero no la forma facial moderna o el tamaño relativo del cerebro. Luego, el Homo sapiens arcaico mostró una mezcla de características que aparecieron en diferentes momentos y lugares, con un conjunto completo de características modernas apareciendo hace aproximadamente 100.000 años.
Si se intenta decidir "quién engendró a quién" y establecer una filogenia precisa, o deducir las causas evolutivas inmediatas para el origen y la propagación de características particulares, entonces más fósiles ayudarían a resolver muchas de las preguntas específicas... a eso se referían Gee, Lewin, Holden y Gould. Los científicos quieren discernir el "quién, cuándo, dónde y cómo" de la evolución, no solo el "si".
McKee:
Falta: Enlace
De lo muy poco que se encuentra, la conclusión básica es que hay básicamente 2 tipos de fósiles de homínidos: los del género Homo y los del género Australopithecus. Todo lo del género Homo (Homo erectus, habilis, neanderthalensis, sapiens, etc.) tiene cráneos y formas corporales que son muy similares a los humanos modernos--a menudo hasta el punto de que podrían estar dentro del posible rango de variación genética humana moderna.
Como se mencionó anteriormente, Homo habilis tenía proporciones de extremidades similares a las de los simios, pero la morfología de un bípedo. Mientras que muchas de las características individuales de los cráneos de los miembros posteriores del género Homo pueden estar dentro del rango humano moderno, el conjunto completo es notablemente diferente. Por ejemplo, se hace mucho énfasis en la observación de que la capacidad craneal de Homo erectus está dentro del rango humano moderno. Bien, pero la relación del tamaño del cerebro con el tamaño del cuerpo está fuera del rango humano moderno.
McKee:
Todo lo demás del género Australopithecus (del cual se deriva el famoso fósil "Lucy"), se parece mucho más a un chimpancé (Australopithecus significa "mono del sur"). Existen formas "robustas" de Australopithecus que no se parecen mucho a los chimpancés, pero estas tampoco se parecen absolutamente nada a Homo, y los evolucionistas no creen que estén en la línea humana.
De hecho, Australopithecus SÍ parece similar al chimpancé en muchos aspectos. También tiene un tamaño relativo del cerebro, dientes, huesos y postura y marcha inferidos, más derivados hacia humanos que hacia chimpancés ... exactamente como uno predeciría de un miembro de la línea homínida que evolucionó poco después de la divergencia de un ancestro común de las líneas que llevan a humanos y chimpancés modernos.
McKee:
¿Qué fósiles hay que vinculen a los miembros similares a los chimpancés de Australopithecus con los fósiles del género Homo, que básicamente representa a los humanos modernos? La respuesta es nada. Entre ambos existen grandes diferencias esqueléticas en cuanto a la locomoción49, 51 y una gran brecha en el tamaño del cerebro50, sin nada en medio. El "eslabón perdido", parece, sigue perdido.
La naturaleza intermedia del esqueleto postcraneal de Homo habilis, como se mencionó anteriormente, contradice la afirmación anterior sobre la locomoción. Pero ¿dónde está el "gran vacío" en el tamaño del cerebro? Por ejemplo, Australopithecus africanus tiene una capacidad craneal promedio de 440 cc (centímetros cúbicos), mientras que el promedio para Homo habilis es de 610 cc, y sus rangos se superponen. No hay un gran salto allí. Sin embargo, más útil es una medida del tamaño relativo del cerebro: el Cociente de Encefalización, o EQ. Básicamente, el EQ nos indica cuánto más de cerebro tiene un animal en comparación con un mamífero típico de su tamaño corporal. Así, por ejemplo, un chimpancé tiene un EQ de 2.0: el doble del cerebro esperado para un mamífero del tamaño de un chimpancé. Ahora, veamos los promedios de EQ para los principales taxones fósiles de homínidos y busquemos grandes vacíos: Australopithecus afarensis: EQ=2.2; A. africanus: EQ=2.5 (por cierto, considerablemente más que un chimpancé moderno); Homo habilis/rudolfensis: EQ=3.1; Homo erectus temprano: EQ=3.3; Homo erectus tardío: EQ=4.0; Homo sapiens moderno: EQ=5.8.
¿El "eslabón perdido" sigue perdido? Sí, ya que cada vez que los paleoantropólogos encuentran un eslabón fósil más de la cadena, los creacionistas encuentran dos eslabones más perdidos (es decir, entre el eslabón y su antecesor, y entre el eslabón y su descendiente). A la actual tasa de descubrimiento y publicación de homínidos fósiles, el número de eslabones perdidos debería aumentar en un orden de magnitud para 2050.
Después de proporcionar citas de varias autoridades, incluyendo a Gee, que señalan la falta de fósiles de homínidos y la naturaleza subjetiva de los muchos árboles filogenéticos de la evolución humana, Wells está justificado en preguntar: «[d]ada la naturaleza altamente subjetiva de la paleoantropología—como reconocen sus propios practicantes—, ¿qué puede decirnos el campo de manera fiable sobre el origen humano?» (Íconos, pág. 223). La respuesta, parece, es definitivamente que no hemos evolucionado sin duda de los simios.
[Nota: la siguiente respuesta a la sección "Missing: Link" de Luskin fue escrita antes de que se proporcionara la respuesta de McKee.]
Luskin afirma que los homínidos fósiles pueden dividirse en dos grupos: los Australopithecus similares a los chimpancés y los Homo, en los que incluye al Homo habilis, diciendo que tienen "cráneos y formas corporales que son muy similares a los humanos modernos". Esto es erróneo de manera asombrosa. Si bien es cierto que los australopitecinos tenían muchas características corporales similares a las de los simios, ¿cuántos chimpancés caminan bípedamente (sobre dos piernas) como lo hacían los australopitecinos? Y si bien la evidencia postcraneal (es decir, no parte del cráneo) para el Homo habilis es escasa, es claro a partir de la evidencia fósil (en particular OH 62) que tenía una forma corporal similar a la de los australopitecinos. Hoy en día, lo que históricamente se clasificó como homo habilis, los habilines, se ha dividido en dos especies, H. habilis y H. rudolfensis, por muchos investigadores. El H. habilis incluye un cráneo que ningún creacionista especialista considera humano, mientras que el H. rudolfensis tiene un cráneo que algunos creacionistas piensan que es de un simio y otros que es de un humano, y sin embargo los dos son muy similares. Recientemente, Bernard Wood, un investigador influyente, y Mark Collard publicaron "The Human Genus" en Science (vol. 284, pp. 65-71, 2 de abril de 1999), en el que propusieron que ambas estas especies habiline sean reclasificadas como Australopithecus y no como Homo. Varios otros investigadores han propuesto previamente que algunos fósiles habiline deberían considerarse australopitecinos. Y Luskin quiere que creamos que hay una línea clara entre Homo y Australopithecus? Pero dejaré que los lectores juzguen por sí mismos. La siguiente tabla tiene algunas imágenes de algunos cráneos de Homo ergaster que tenían una forma corporal bastante moderna, las dos especies de habilines y una especie de australopitecino. Los enlaces debajo de las imágenes llevan a una foto más grande o a más información sobre el fósil en particular. Las capacidades craneales estimadas (tamaños cerebrales) de cada especie se toman del artículo de Wood y Collard.
| Especie | Capacidad Craniana |
FH = Preguntas frecuentes sobre homínidos fósiles SI = Programa de Orígenes Humanos del Instituto Smithsonian |
||
|---|---|---|---|---|
| Homo Ergaster | 854 cm3 |  FH SI |
 FH SI |
|
| habilines | Homo (o Australopithecus) rudolfensis | 752 cm3 | 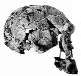 FH SI |
|
| Homo (o Australopithecus) habilis | 552 cm3 | 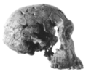 FH SI |
 FH SI |
|
| Australopithecus africanus | 457 cm3 |  FH SI |
 SI |
|
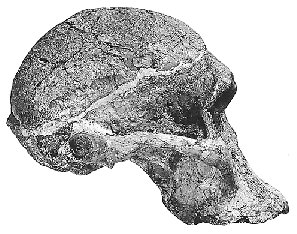{kind=link}
Existen muchos otros fósiles también. El lector es invitado a ir a este enlace y tratar de trazar la línea entre "mono" y "humano". No es un asunto sencillo como uno esperaría si los monos y los humanos fueran creaciones separadas, como lo demostró la tabla de opiniones de los creacionistas de Jim Foley sobre qué son los homínidos fósiles.
-- Mike Hopkins
Conclusión:
Wells no es un fraude
Aparte de sus dudas sobre el darwinismo, una revisión de las acusaciones de Tamzek contra Wells revela que no hay un solo caso establecido donde Wells esté equivocado, y pocos donde incluso esté claramente en desacuerdo con el pensamiento científico mainstream. El único error bona-fide en Icons podría encontrarse en la figura 5-3 de Wells sobre la explosión cámbrica, la cual Tamzek no verifica, y este supuesto error en sí mismo no afectaría la fuerza de los argumentos de Wells sobre la explosión cámbrica. ¿Cómo es que Wells puede parecer estar en acuerdo con la ciencia mainstream en casi todos sus argumentos (incluso Eugenie Scott dice que Wells es "usualmente técnicamente correcto"89), lo cual debería tener un efecto profundamente negativo sobre la evidencia para la teoría de la evolución, y sin embargo Wells difiere de la ciencia mainstream en su conclusión de que la evolución podría ser falsa? La respuesta más probable yace en la interpretación de Wells de los datos que recopila al analizar el nivel de errores en los libros de texto.
El texto completo de la reseña de Eugenie Scott se puede encontrar en:
http://www.scienceormyth.org/icons%20of%20evolution.html
Luskin cita sus palabras fuera de contexto, como se señaló aquí. Esto es un poco más de lo que ella escribió:
...Esto se debe a que Wells presenta una visión sistemáticamente engañosa de la evolución. Las oraciones individuales en Icons son usualmente técnicamente correctas, pero están artísticamente enlazadas para llevar al lector fuera del camino de la biología evolutiva real y hacia un seto de malentendidos.... [Énfasis añadido.]
-- Mike Hopkins
Dése en la luz
En conclusión, Tamzek cita a Teodosio Dobzhansky, un evolucionista teísta, para demostrar que se puede creer en Dios y en la evolución. En la cita que proporciona Tamzek, Dobzhansky parece estar argumentando únicamente contra una perspectiva de la Tierra joven, lo cual no es en absoluto lo que Wells defiende. De hecho, Wells podría incluso estar de acuerdo con Dobzhansky en que los textos religiosos no son "libros de texto de astronomía, geología, biología y antropología". Donde Wells discreparía es en que, por lo tanto, la conclusión debería ser que, desde un punto de vista empírico, Dios no tuvo un efecto directo sobre el proceso de creación, y que Dios creó todo a través de una evolución no dirigida. La Escritura bíblica puede decir poco sobre los detalles de los procesos reales que Dios utilizó para crear, sin embargo, es claro que Dios quiso que las cosas fueran como son57. A Dobzhansky le resultaría difícil encajar la aleatoriedad de la evolución en la finalidad de Dios. Para el teísta dispuesto a arriesgar su credibilidad académica reconociendo las contrapruebas a la vaca sagrada de la biología105, estos problemas pueden resolverse. Si Dobzhansky hubiera vivido para ver el nacimiento del movimiento del diseño inteligente, quizás él también, como teísta, habría abrazado su enfoque más abierto.
A un nivel fundamental, todos los creacionistas creen generalmente en el diseño inteligente. Sin embargo, de la manera que Él haya elegido hacerlo, todos los creacionistas creen que Dios creó el mundo y sus especies, incluido el hombre, de manera intencional. En su ensayo, "Nothing in Biology Makes Sense Except in the Light of Evolution", Dobzhansky se autodenominó explícitamente un creacionista y un evolucionista, lo que por sustitución significa que se consideraba un defensor del diseño inteligente evolutivo o de la evolución teísta. (De ahora en adelante, todas las referencias a "diseño inteligente" se entenderán como referencias al "diseño inteligente anti-evolucionista" y no a la evolución teísta. No obstante, debe enfatizarse que, en general, no existe necesidad de que una inteligencia utilice procesos estrictamente no darwinianos para crear las diversas formas de vida en este mundo. La evolución teísta es una posición legítima del diseño inteligente y, notablemente, es solo la versión de Wells o de su clase del diseño inteligente anti-evolucionista la que convierte a la evolución o a los métodos de la ciencia en enemigos). Por lo tanto, intentar apoyar la idea de que Dobzhansky abogaría por cualquier forma de un creacionismo anti-evolucionista, especialmente las formulaciones modernas del diseño inteligente, sería un desafío considerable.
En su respuesta a Tamzek, Luskin intenta lograr precisamente este hazaña hercúlea. Luskin intenta argumentar que Dobzhansky era ignorante tanto de las contribuciones del azar aleatorio a la evolución como de los métodos modernos de diseño inteligente para explicar las observaciones. Si esto fuera cierto, Luskin razona que Dobzhansky podría haber apoyado el razonamiento moderno del diseño inteligente si simplemente se le hubiera dado la oportunidad. Desafortunadamente, estas afirmaciones no son ciertas y la afirmación de Luskin sobre un posible apoyo de Dobzhansky es improbable.
Primero, el movimiento del diseño inteligente no es nuevo. Sus raíces modernas se remontan al menos a Michael Polanyi, quien fue contemporáneo de Dobzhansky. Incluso omitiendo las contribuciones modernas de Polanyi a la literatura del diseño inteligente, el "Argumento del Diseño" del Reverendo William Paley describió por primera vez el argumento básico del diseño inteligente, y Paley vivió mucho antes incluso que Darwin.
A un nivel básico, los argumentos del diseño inteligente sostienen que un fenómeno determinado no puede explicarse mediante la ciencia y, por lo tanto, requieren la existencia de Dios. [2] Las contribuciones de Behe y Dembski a la literatura del diseño inteligente, aunque suenen modernas y sean esotéricas, aún se basan en la misma lógica defectuosa de Paley; solo el fenómeno en cuestión ha cambiado con el tiempo. Por lo tanto, los argumentos del diseño inteligente no han cambiado de manera significativa desde que Paley los formuló por primera vez. Dado que Dobzhansky no apoyó la razonamiento de Paley, es poco probable que hubiera apoyado también la razonamiento de Behe o Dembski.
Existe también una alta probabilidad de que los argumentos modernos de diseño inteligente estén diseñados para escurrirse por la cláusula de establecimiento de nuestra Constitución. De hecho, los defensores modernos del diseño inteligente solo han eliminado las afirmaciones positivas sobre el diseñador que hizo Paley: donde Paley argumentó explícitamente por Dios, los defensores modernos del diseño inteligente argumentan por un "diseñador" más vago y oficialmente sin nombre. Tal prestidigitación argumentativa no altera realmente la lógica del argumento de Paley. Por el contrario, simplemente hace poco clara la identidad del diseñador, los propósitos del diseño del diseñador y otras afirmaciones positivas sobre el diseño. De nuevo, el argumento de los defensores modernos del diseño inteligente no es sustancialmente diferente del argumento de los defensores premodernos del diseño inteligente.
Que Luskin afirme que Dobzhansky nunca tuvo la oportunidad de considerar métodos o argumentos de diseño inteligente es negar tanto la larga historia del creacionismo de diseño inteligente como la congruencia entre los argumentos modernos y premodernos de diseño inteligente. En su lugar, es probable que Dobzhansky los incluyera presagiantemente incluso mientras hacía el caso contra los creacionismos anti-evolución en su ensayo que precedió tanto a Behe como a Dembski.
En segundo lugar, la evolución implica, al menos en parte, una contribución del azar aleatorio -- siempre lo ha hecho, desde que Darwin escribió su primera edición de Origen. Por lo tanto, como científico contribuyente al campo de la biología y al estudio de la evolución, es poco probable que Dobzhansky no hubiera considerado el azar en la evolución que consideraba perfectamente compatible con la teología moderna. Considere la siguiente cita de su ensayo.
La diversidad orgánica se vuelve, sin embargo, razonable y comprensible si el Creador ha creado el mundo vivo no por capricho sino por evolución impulsada por la selección natural. Es incorrecto considerar la creación y la evolución como alternativas mutuamente excluyentes. Soy creacionista y evolucionista. La evolución es el método de creación de Dios, o de la Naturaleza. La creación no es un evento que ocurrió en el año 4004 a. C.; es un proceso que comenzó hace aproximadamente 10 mil millones de años y que aún está en curso.
Una sección entera de este ensayo está dedicada al análisis de la diversidad orgánica. No hay duda de que Dobzhansky atribuyó esa diversidad al azar, al menos en parte. Sin embargo, las palabras "aleatorio" o "azar" no aparecen en el ensayo de Dobzhansky. No obstante, en otros escritos queda muy claro que la evolución implica contribuciones del azar y la casualidad. Considere esta cita del libro de texto de Futuyma:
Como dijo Dobzhansky (1970), "Puede parecer una lamentable imperfección de la naturaleza que la mutabilidad no esté restringida a los cambios que mejoran la adaptabilidad de sus portadores. Sin embargo, solo un vitalista Pangloss podría imaginar que los genes saben cómo y cuándo es bueno para ellos mutar." {Futuyma, Biología Evolutiva 2ª ed. 1986, p. 76}
O considera esta cita del American Scientist:
El proceso de mutación es la única fuente conocida de nuevos materiales de variabilidad genética, y por tanto de la evolución. {T. Dobzhansky, American Scientist, v. 45, 1957, p.385}
A la luz de estas declaraciones, parece que Dobzhansky reconoció efectivamente el azar en su comprensión de la evolución. "Nothing in Biology Makes Sense Except in the Light of Evolution" fue escrito en 1973 y las citas anteriores fueron escritas en 1970 y 1957. Por cualquier evaluación razonable, Dobzhansky incluyó influencias aleatorias en la evolución que consideraba compatible con el teísmo.
Luskin sugiere que Dobzhansky era ignorante sobre la aleatoriedad en la evolución, aunque Dobzhansky sabía muy bien de la aleatoriedad involucrada en la evolución. Luskin argumenta que Dobzhansky era ignorante sobre los argumentos modernos de diseño inteligente, aunque Dobzhansky sabía muy bien las falacias en la versión prototípica de Paley del mismo. La sugerencia de Luskin de que Dobzhansky podría haber estado de acuerdo con el creacionismo de diseño inteligente es, por lo tanto, no respaldada y, de hecho, altamente improbable.
A diferencia de Tamzek, Wells utiliza las citas de Dobzhansky con fines puramente científicos: para demostrar que la famosa cita de Dobzhansky—"[n]ada en biología tiene sentido más que a la luz de la evolución"104—representa un alejamiento del método científico, ya que presupone que todo en biología debe encajar bajo el paradigma evolutivo:
"[L]a ciencia en su mejor versión persigue la verdad. Dobzhansky estaba completamente equivocado, y lo mismo ocurre con quienes continúan recitando su mantra anticientífico. Para un verdadero científico, nada en biología tiene sentido más que a la luz de la evidencia." (Íconos, pág. 248)
Dobzhansky escribió un ensayo titulado "Nada en biología tiene sentido a menos que se vea a la luz de la evolución". Notablemente, Dobzhansky escribió un ensayo con este título; no escribió solo el título. En el ensayo, Dobzhansky aclara por qué tituló su ensayo de esa manera: la evolución explica las observaciones mejor que cualquier otra explicación ofrecida y la biología sin evolución sería un montón de observaciones sin un tema unificador.
Considere los siguientes extractos del ensayo en cuestión:
El entorno no impone cambios evolutivos a sus habitantes, como postulaban las ahora abandonadas teorías neolamarckianas.
Todo esto es comprensible a la luz de la teoría de la evolución; pero, ¿qué operación tan absurda habría sido, por parte de Dios, fabricar una multitud de especies ex nihilo y luego dejar que la mayoría de ellas se extinguiera!
Se imaginan que todas las especies existentes fueron generadas por un decreto sobrenatural hace unos pocos miles de años, casi tal como las encontramos hoy. Pero ¿qué sentido tiene tener hasta 2 o 3 millones de especies viviendo en la Tierra?
¿Estaba el Creador de buen humor cuando creó Psilopa petrolei para los campos petrolíferos de California y especies de Drosophila para vivir exclusivamente en algunas partes del cuerpo de ciertos cangrejos terrestres en solo ciertas islas del Caribe?
¿Y si no hubo evolución y cada una de las millones de especies fue creada por un mandato separado? Sin embargo ofensiva que la idea pueda ser para el sentimiento religioso y para la razón, los antievolutionistas deben acusar nuevamente al Creador de estafar. Deben insistir en que Él dispuso deliberadamente las cosas exactamente como si su método de creación fuera la evolución, con la intención de engañar a los buscadores sinceros de la verdad.
Observe cómo Dobzhansky demuestra específicamente cómo ha considerado otras explicaciones, tanto no naturales como naturales, pero las encontró todas insuficientes. Tras haber considerado esas alternativas, que probablemente son representativas de todas las formas de creacionismo anti-evolucionista o evolución no neodarwiniana (evolución lamarckiana, etc.), Dobzhansky tituló su ensayo como lo hizo. Si Dobzhansky no hubiera considerado ninguna alternativa, la objeción de Luskin sería precisa: existe la posibilidad de que otra hipótesis de biodiversificación encaje con los datos; por lo tanto, considerar la evolución como la única explicación posible es o bien totalmente inválido o dogmático.
Desafortunadamente para Luskin, el ensayo deja muy claro que el título no transmite ni desarrolla completamente las ideas de Dobzhansky. Es posible explicar los datos desde una perspectiva no evolutiva, pero como Dobzhansky señala a menudo en el ensayo, esas perspectivas son absurdas y no son tan sólidas ni tan útiles como la evolución. Por lo tanto, el título de Dobzhansky sigue siendo válido y el punto declarado por Luskin puede ser fácilmente descartado.
Existe otra manera en que la objeción de Luskin podría ser válida, aunque él no la formula por sí mismo. ¿Qué pasaría si la evolución fuera fácilmente eliminada del campo de la biología en bloque? Si los científicos pudieran de alguna manera comprender la biología sin la evolución, entonces la sugerencia implícita en una lectura literal del título de Dobzhansky —de que nada en la biología podría tener sentido sin la evolución— sería inexacta y la objeción de Luskin podría sostenerse.
Lo más probable es que fuera a esta misma objeción a la que Dobzhansky escribió en su ensayo:
Visto a la luz de la evolución, la biología es, quizás, la ciencia más intelectualmente satisfactoria e inspiradora. Sin esa luz, se convierte en un montón de hechos diversos, algunos interesantes o curiosos, pero que no forman una imagen significativa en su conjunto.
Dobzhansky está diciendo que la separación de la evolución de nuestra comprensión moderna de la biología sería un proceso difícil o haría que la biología fuera incomprensible por completo. En cualquier otro lenguaje, está diciendo que nada en la biología tiene sentido excepto en la luz organizadora y explicadora de la evolución, una interpretación justificable que también socava la objeción de Luskin.
Los creacionistas a menudo intentan socavar la lógica del ensayo de Dobzhansky alegando que efectivamente existen observaciones en biología que no requieren una perspectiva evolutiva para ser comprendidas. Wells parece seguir este camino al argumentar también contra el título en lugar del ensayo, aunque prefiere atacar desechando la evolución como un mero paradigma. Ambos argumentos presuponen que el lector ha leído solo el título y no el ensayo que lo lleva. Si se lee el ensayo de Dobzhansky, la elección del título está justificada, incluso si es un resumen torpe del propio ensayo. Si Luskin argumenta que el título del ensayo de Dobzhansky no desarrolla el ensayo de Dobzhansky tan completamente como lo hace el propio ensayo, su argumento es poco notable y fácilmente desechable al recordar a los lectores que la comprensión de los temas rara vez se logra juzgando los proverbiales libros por su portada.
En realidad, los puntos de Luskin y Wells pueden abordarse fácilmente leyendo el ensayo en cuestión [http://www.2think.org/dobzhansky.shtml]. El título de ese ensayo está completamente justificado por el desarrollo del mismo: no representa una desviación del método científico, ni presenta defectos en su razonamiento.
En cuanto a los métodos de la ciencia, ¿qué ironía que Wells, como creacionista del diseño inteligente, acusara a Dobzhansky de haberse desviado del método científico. Las respuestas a las preguntas biológicas que los defensores del diseño inteligente proponen, porque esas respuestas invocan portadores de cambios naturales "sobrenaturales" inobservables e inmedibles, son respuestas que no pueden ser puestas a prueba por la ciencia. Son los creacionistas del diseño inteligente, y no Dobzhansky, quienes se desvían del método científico —y se desvían de él de manera general—. Por lo tanto, Wells no puede acusar seriamente a nadie de desviarse de los métodos de la ciencia sin incurrir en una flagrante hipocresía.
El punto de Wells es que la evidencia, no un paradigma, debería controlar nuestra comprensión de los datos. Después de examinar las debilidades en las líneas de evidencia más comunes utilizadas para apoyar la teoría de la evolución, al menos dos cosas son claras: la evidencia a favor de la teoría de la evolución seguramente falta en algunos lugares importantes, y Íconos de la Evolución será una verdadera molestia para los evolucionistas durante los próximos años.
La comprensión científica actual del mundo "natural" observable y medible es la que explica mejor las observaciones y predice con mayor precisión nuevos datos. Los nuevos datos y las nuevas teorías se consideran a la luz de estas comprensiones actuales. Es el proceso de la ciencia reevaluar continuamente el estado de las explicaciones consideradas, manteniendo las teorías que mejor explican y predicen los datos y desvinculando o rechazando directamente el resto. Al definir la evolución como la comprensión científica actual de la biodiversificación, se debe inferir que, por lo tanto, explica y predice mejor los hallazgos del mundo comprobable.
Wells reconoce que la evolución tiene un marco teórico amplio al cual se aplican nuevos datos. Sin embargo, la descalifica como meramente un "paradigma", como si la evolución fuera una teoría que, en principio, no podría ser refutada por observaciones naturales. Al hacer este argumento, ignora el hecho de que desde el día en que Darwin concibió la evolución, esta fue capaz de ser refutada. Hasta ahora, no ha sido así.
No es un tratamiento sesgado contra la evolución o a favor del diseño inteligente lo que mantiene a la evolución como la comprensión científica actual de la biodiversidad y otras observaciones. Lejos de ser simplemente un "paradigma" para guiar el pensamiento en biología o no, la evolución es en cambio el caballo de batalla de una teoría científica. Ha guiado innumerables experimentos, ha explicado tantas observaciones y mediciones en diversos campos y ha sido verificada y reforzada tan completamente que está tan cerca de ser un hecho como la ciencia probablemente pueda llegar a estar. Es verdaderamente "La teoría integradora de la biología."
Por lo tanto, la sugerencia de Wells de que la evolución es simplemente un "paradigma" favorecido o carece de un respaldo teórico útil es completamente incorrecta. La evolución es, en cambio, la comprensión científica actual porque la evidencia la ha respaldado robustamente.
En cuanto a la evidencia, Wells a menudo se ha presentado a sí mismo como un biólogo que simplemente discrepa con la evolución. Si Wells realmente actuara como un científico o utilizara realmente la formación que aprendió en su búsqueda de un título avanzado en biología, estaría trabajando para desarrollar la evidencia necesaria para apoyar su caso contra la evolución. En cambio, Wells argumenta contra la definición de ciencia, calificando de sesgada la restricción de la ciencia a hipótesis "naturales" comprobables. Wells probablemente reconoce la naturaleza robusta de la evolución y, con el fin de promover su admitida cruzada contra el darwinismo por la cual buscó acreditación avanzada como biólogo en primer lugar, considera más fácil socavar los métodos de la ciencia que apoyar científicamente su hipótesis.
En lugar de sucumbir al estratagema de Wells, el lector es invitado a examinar la evidencia que respalda tanto la evolución como el diseño inteligente y tomar una decisión como lo haría un científico. La revista Evolución tiene más de 18 artículos revisados por pares rigurosamente cada mes. En todos los periódicos, se publican varios artículos de investigación original sobre la evolución todos los días, y son posibles innumerables estudios adicionales solo gracias a los conocimientos que proporciona la evolución o a las predicciones que hace. Por el contrario, es realmente notable que el diseño inteligente no tenga tales artículos. Ni siquiera intentan hacer investigación real porque no pueden; ningún dato puede contradecir jamás a un agente sobrenatural postulado y apelar a las acciones inobservables e inmedibles de agentes sobrenaturales para explicar datos naturales simplemente no es una hipótesis falsable en la ciencia.
A pesar de la ausencia completa de evidencia científicamente significativa, Wells y los otros creacionistas del diseño inteligente solicitan que la "evidencia" del diseño inteligente sea permitida junto a la evolución en los salones de clases de ciencias, basándose en que la evidencia y no un "paradigma" debería controlar nuestra comprensión de los datos. Si uno realmente basara la inclusión o exclusión de las discusiones en los salones de clases de ciencias sobre la evidencia, entonces el diseño inteligente simplemente no aparecería en un salón de clases de ciencias. Por lo tanto, el argumento de Wells se vuelve tan hipócrita como su anterior: no tiene evidencia para respaldar su afirmación más allá de un apelo a la ignorancia.
Luskin, apelando a la investigación de Wells, afirma entonces que existen "defectos" en la teoría de la evolución. Su descripción peyorativa de la investigación actual como "defectos" es sin duda digna de reproche, el cual se ofrece a continuación. En primer lugar, esta respuesta debe comenzar recordando a los lectores la sustancia de la crítica original de Tamzek a Wells. Cualquier investigador imparcial reconocerá que Wells no puede ser confiado para transmitir adecuadamente el estado de la evolución a los lectores. Simplemente porque Wells considera que ciertas líneas de evidencia a favor de la evolución son "defectuosas" no significa que observadores abiertos a la mente las considerarían defectuosas. Luskin haría bien en buscar otro apoyo para sus conclusiones que las declaraciones en el libro de Wells.
Es lamentable, pero revelador, que Luskin describa peyorativamente las áreas activas de investigación en evolución como "defectos". Al hacerlo, intencionalmente connota la ilegitimidad de la evolución incluso mientras esta avanza a través de una gloriosa era de éxito y utilidad. De hecho, el uso peyorativo de Luskin revela más sobre su sesgo personal contra la evolución que sobre la evolución misma. Claramente, existe la necesidad de más investigación en el campo de la evolución; nadie duda de esto. Sin embargo, se necesita más investigación en la evolución por las mismas razones por las que se necesita más investigación en la química. Es decir, siempre habrá más por aprender sobre la química y algunas de las cosas que creemos saber sobre la química resultarán ser incorrectas; sin embargo, esto no apoya la alquimia.
El hecho es que la ciencia es una disciplina activa y existe y siempre habrá un horizonte más allá del cual aún no tenemos comprensión y en el cual nuestra comprensión es incompleta. En y cerca de este horizonte, nuestros modelos están sujetos a modificación, quizás rechazo, a medida que se hace disponible nueva información. Sin embargo, la existencia de incertidumbre en este horizonte no niega nuestro conocimiento en el primer plano bien transitado. [1] Con disculpas a Einstein, "Si supiéramos todo sobre [la evolución], no sería ciencia."
Es cierto que aún se necesita mucha investigación sobre la evolución: los detalles sobre los mecanismos de la evolución, las rutas particulares que tomó en linajes específicos, la aplicación de la evolución a otros problemas o datos no necesariamente relacionados con la especiación, etc., todo esto necesita ser investigado y resuelto. Sin embargo, la intención de la declaración de Luskin —que existen "defectos importantes" de manera pivotante en la teoría de la evolución y que estos "defectos" socavan la evolución en su conjunto— es inválida por al menos dos razones.
En primer lugar, es importante comprender que en el "primer plano bien transitado" de los fundamentos de la evolución, este está sólidamente articulado con un apoyo científico adecuado —o mejor dicho, abrumador—. El debate sobre la evolución o la investigación realizada sobre la evolución no versa sobre si ocurrió, sino sobre cómo ocurrió. Puesto en un lenguaje diferente, en el nivel educativo en el que los conceptos evolutivos se investigan activamente, uno probablemente estaría inscrito en un curso de posgrado: la evolución a nivel de secundaria y pregrado está tan cerca de lo factual como la ciencia puede llegar a estar. Simplemente, las áreas de investigación activa en la evolución —las supuestas "deficiencias" de Luskin— no son de un tipo que haría dudar al imparcial de si la evolución ocurrió; los "frentes" de la evolución se encuentran muy lejos de los fundamentos establecidos, sobre los cuales no existe ninguna objeción científica seria. Por lo tanto, Luskin está muy equivocado al afirmar que hay "deficiencias" en la evolución, si su intención al citar esas "deficiencias" es sugerir que estas socavan la evolución.
En segundo lugar, observa la suposición no expresada de Luskin: el fracaso de nuestra comprensión actual de la evolución puede ser tomado como un éxito para el diseño inteligente. Esta lógica sugiere que el diseño inteligente es una explicación por defecto: descartar la evolución y el diseño inteligente es lo único que queda. Su suposición es seriamente defectuosa por al menos dos razones. Primero, un fallo común en un argumento desde la ignorancia es que superar la ignorancia en cuestión fácilmente lo debilita. En general, dada la historia de la ciencia y su gloriosa historia de superar la ignorancia de los fenómenos, las lagunas en nuestra comprensión actual de la evolución serán llenas eventualmente. Específicamente, el intento de Luskin de descartar las comprensiones actuales de la evolución como "defectuosas" no hace mucho para descartar las comprensiones futuras de la evolución.
Segundo y más importante, ¿qué hay del diseño inteligente que hace que Luskin piense que merece un estatus por defecto? La comprensión de la evolución que tenemos actualmente —que tiene áreas activas de investigación en conceptos evolutivos avanzados— tiene un marco teórico, predictivo, explicativo y útil mucho más amplio que el creacionismo del diseño inteligente, el cual, francamente, no tiene ninguna de estas cosas. Considerar que "la evidencia para la teoría de la evolución falta seguramente en algunos lugares importantes" y concluir que el diseño inteligente es por lo tanto una mejor explicación para observaciones perfectamente naturales es abandonar la perspectiva, la razón o los métodos de la ciencia.
En cuanto al libro de Wells ser un "dolor de culo para los evolucionistas", efectivamente ha asumido el papel de una mosca socrática, despertando respuestas en las comunidades científicas y religiosas dormidas. Anteriormente, estas comunidades no consideraban el creacionismo como una amenaza. Tras la publicación del libro de Wells, se volvieron más activas en abordar el creacionismo.
En consecuencia, en cualquier lugar donde Wells o los creacionistas del diseño inteligente hablen ahora, los científicos y los teólogos refutan su retórica con un interés y pasión que no habrían tenido de otro modo. Esta refutación de Luskin, por ejemplo, ha sido creada con la asistencia de muchos colaboradores, algunos de los cuales son profesionales del campo que no suelen participar en debates creacionistas. En Kansas, y probablemente en otros estados también, existe un interés en la evolución que no estaba presente antes de la publicación de Íconos. Por cualquier medida, el libro de Wells ha hecho más por promover la evolución que por dañarla.
El lector es recordado nuevamente de la refutación original de Tamzek a Wells, en la que se expone la flagrante tergiversación de la ciencia por parte de Wells. Dado que Wells ha fallado en demostrar integridad académica, la publicación de su libro conlleva o conllevará un gran costo personal: Wells no será considerado un investigador o autor reputado por ningún grupo distinto de los fieles del diseño inteligente.
Considerando las bendiciones para la evolución paradójicamente proporcionadas por Íconos y los costos para la credibilidad de Wells, si la publicación de Íconos como un "dolor en el trasero para los evolucionistas" es una victoria para Wells, es tanto una victoria pírrica para él como una que tanto la ciencia como la religión están dispuestas a conceder.
Notas finales:
#1: Abell, GO (1983). "Las edades de la Tierra y el Universo." Los científicos se enfrentan al creacionismo: Nueva York: WW Norton & Company.
#2: Pennock, Robert T. (2000) Torre de Babel: La evidencia contra el nuevo creacionismo. Cambridge: MIT Press. P. 163.
-- Burt Humburg
Nota: Este documento fue editado por varias personas, incluyendo a Nic Tamzek y Mike Hopkins. La introducción original fue escrita por Nic, aunque varias otras personas han realizado cambios en ella.